ポイント
- 脳での特異的な発現はGnT-IX遺伝子周辺のヒストンの活性化に依存
- ヒストンの活性化がGnT-IX遺伝子発現を促す転写因子を呼び込む
- 臓器、組織、細胞で糖鎖の構造が異なる理由の解明に手掛かり
要旨
独立行政法人理化学研究所(野依良治理事長)は、糖転移酵素※1の1つ「GnT-IX」の遺伝子「GnT-IX遺伝子」が脳だけで発現する仕組みの解明に挑み、この遺伝子近傍に存在するヒストン※2の活性化が重要であることを発見しました。これは、理研基幹研究所(玉尾皓平所長)システム糖鎖生物学研究グループ(谷口直之グループディレクター)疾患糖鎖研究チームの木塚康彦特別研究員、北爪しのぶ副チームリーダー、ケミカルゲノミクス研究グループの吉田稔グループディレクターらの研究成果です。
糖鎖とは、グルコース(ブドウ糖)などの糖が鎖状につながった分子で、タンパク質や脂質の上にさまざまな形で付加しており、その機能を調節しています。糖鎖の付加や構造に異常があると、筋ジストロフィーや糖尿病、慢性閉塞性肺疾患、がん転移などの発症につながる可能性があることが分かってきています。糖鎖は、細胞の中で糖転移酵素と呼ばれる一群の酵素によって作られますが、その分布は動物体内で均一ではなく、各臓器、組織、細胞によってそれぞれ異なる形で作られます。これまで、各臓器、組織、細胞によってなぜ糖鎖の構造が異なるのかは分っていませんでした。
研究グループは、2003年に発見した糖転移酵素の1つGnT-IXがなぜ脳にだけ存在するのかに着目しました。このGnT-IXは、筋ジストロフィーの原因となるO-マンノース型糖鎖※3の合成にも関わっていることが知られています。まず、マウスの脳の細胞と腎臓や肝臓の細胞を用いて、クロマチン免疫沈降法※4を実施した結果、ヒストンの活性化(翻訳後修飾※5)がGnT-IX遺伝子の発現を制御していることを明らかにしました。また、ゲルシフト法※6などにより、GnT-IX遺伝子の発現を制御する2つの転写因子※7、CTCFとNeuroD1を同定しました。さらに、これらの転写因子はヒストンが活性化されることによってGnT-IX遺伝子の近くに集まることも発見しました。これらの結果から、ヒストンの活性化、転写因子の呼び込み、GnT-IX遺伝子の発現、という一連の流れが明らかになりました
今回解明したメカニズムは、臓器、組織、細胞それぞれに特異的な糖鎖の構造を形成する根幹である可能性を示しており、糖鎖形成不全が引き起こす疾患発症の原因解明などに役立つと期待されます。
本研究成果は、米国の科学雑誌『The Journal of Biological Chemistry』(9月号)に掲載されるに先立ち、オンライン版(7月19日付け:日本時間7月20日)に掲載されました。
背景
最近、動物体内の半数以上のタンパク質に糖鎖が付加されていることや、付加した糖鎖がそのタンパク質のさまざまな機能を調節していることが分かってきています。糖鎖の形は臓器、組織、細胞によって大きく異なり、これにより固有の機能を発揮しています。糖鎖は、細胞の中の小胞体、ゴルジ体と呼ばれる器官で一連の酵素(糖転移酵素)によって作られますが、各臓器、組織、細胞が持つ酵素の種類は異なっているため、それぞれ特異的な糖鎖が形成されます。つまり、糖転移酵素の元になる遺伝子の発現は、細胞の種類によって調節されています(図1)。しかし、この遺伝子発現の調節の仕組みは明らかにされておらず、どのように特異的な糖鎖が作られるのかも分かっていませんでした。
研究グループは、2003年に谷口直之グループディレクターらが発見した糖転移酵素の1つで、筋ジストロフィーの原因として知られるO-マンノース糖鎖の合成に関与するGnT-IXに着目しました。この酵素は、脳だけに存在するという特徴を持っているため、この酵素の元になるGnT-IX遺伝子は脳だけで発現していることになります。そこで、GnT-IX遺伝子が脳だけで発現している仕組みの解明に挑みました。
研究手法と成果
遺伝子発現は、染色体の構成成分であるヒストンのアセチル化やメチル化といった翻訳後修飾によって調節されることがあります。そこで研究グループは、GnT-IX遺伝子が発現しているマウスの脳の細胞と、発現していない腎臓や肝臓の細胞を用いて、この遺伝子周辺にあるヒストンの活性化の様子を解析しました。ヒストンの特定の部位の修飾を検出する抗体を用いてクロマチン免疫沈降法を行ったところ、GnT-IX遺伝子が発現している細胞だけで、その遺伝子周辺のヒストンがアセチル化などを受けた活性化の状態にあることを確認しました(図2A)。また、ゲルシフト法などにより、GnT-IX遺伝子近傍のDNA領域に結合するタンパク質として、CTCF、NeuroD1という2種類の転写因子を同定することに成功し(図2B)、ヒストンが活性化されている状態ではこの結合が起こりやすいことも見いだしました。これらの結果から、ヒストンが活性化するとCTCFやNeuroD1がGnT-IX遺伝子近傍に結合し、GnT-IX遺伝子の発現が促されることが明らかになりました(図3)。
今後の期待
ヒストンの特定の修飾やDNAのメチル化※8といった、DNA配列に依存しない後天的な染色体の修飾現象はエピジェネティック※9な制御と呼ばれ、遺伝子の調節に重要です。今回の研究により、臓器、組織、細胞に特異的な糖鎖の形成がエピジェネティックな制御を受けていることが明らかになりました。これまで糖鎖の形成に関しては、このような視点からの解析は行われておらず、糖鎖の付加の仕組みを研究する上で大きな発見であるといえます。
今後は、ヒストンが活性化される仕組みを明らかにする必要があります。また、糖鎖の合成に関わる他の遺伝子もGnT-IX遺伝子と同様な仕組みで調節されている可能性があります。これらを明らかにすることで、なぜ糖鎖が臓器、組織、細胞それぞれに特異的に形成されるのかが解明され、糖鎖形成不全によって引きこされる疾患の原因解明などに役立つと期待されます。
発表者
理化学研究所
基幹研究所 ケミカルバイオロジー研究領域
システム糖鎖生物学研究グループ
グループディレクター 谷口 直之(たにぐち なおゆき)
Tel: 048-467-8094 / Fax: 048-467-9616
Tel: 048-467-8104 / Fax: 048-467-9617
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.糖転移酵素
主に細胞の中の小胞体、ゴルジ体と呼ばれる器官に存在し、糖鎖の合成に関わる一群の酵素。活性化された糖供与体を使い、1つ1つ糖を糖鎖に転移していくことで、最終的に成熟した形の糖鎖が作られる。 - 2.ヒストン
染色体の構成成分タンパク質。DNAはヒストンの周囲に巻き付いて存在している。近年、染色体の構成成分としてだけでなく、メチル化やアセチル化といったヒストンの翻訳後修飾が遺伝子発現の制御に重要であることが明らかになりつつある。 - 3.O‐マンノース型糖鎖
タンパク質に近い側の末端の糖がマンノースである型の糖鎖。この糖鎖の形成異常は、ある種の筋ジストロフィーの原因となる。 - 4.クロマチン免疫沈降法
生きている細胞の中でのDNA-タンパク質間の結合を検出する方法。タンパク質を認識する抗体を使ってタンパク質-DNA複合体を沈降させ、タンパク質を除去した後、PCR法によってDNAを検出する。 - 5.翻訳後修飾
タンパク質がRNAから翻訳された後に化学修飾されること。糖鎖付加をはじめ、リン酸化、メチル化、アセチル化などが知られる。 - 6.ゲルシフト法
タンパク質とDNAの結合を検出する方法の1つ。DNAがタンパク質と結合していると分子量が大きくなり、ゲルの中を移動する速度が遅くなる現象を利用している。 - 7.転写因子
特定のDNA配列に結合して遺伝子の発現を制御するタンパク質。 - 8.DNAのメチル化
DNA配列にメチル基(-CH3)が付加されること。特に、CG配列のCのメチル化はよく研究されている。遺伝子の発現を司るプロモーターと呼ばれるDNA領域にはCG配列が多く、この部分がメチル化されると遺伝子の発現が抑制されることが知られている。 - 9.エピジェネティック
DNA配列によらない遺伝子の調節機構のこと。A、T、G、Cの4塩基からなるDNAの塩基配列だけでは説明できない遺伝子発現調節の仕組みとして、近年急速に研究が進んでいる。DNAのメチル化やヒストンの修飾などがこれにあたる。
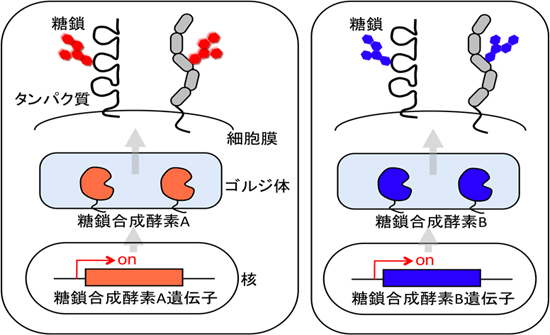
図1 臓器・組織・細胞によって異なる糖鎖合成酵素の発現と糖鎖の形成
異なる細胞では、異なる糖転移酵素のセットが存在している。これは、糖転移酵素の元になる遺伝子の発現が各臓器、細胞、組織ごとに異なるためで、最終的に形成される糖鎖はそれぞれ特異的な形になる。
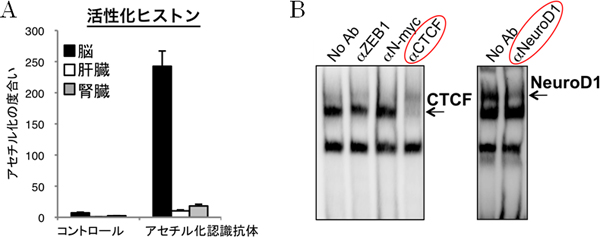
図2 脳特異的なヒストンの活性化と、転写因子CTCF、NeuroD1の同定
- A: マウスの脳、肝臓、腎臓の細胞を用い、ヒストンの活性化の指標となるアセチル化を認 識する抗体を使って、GnT-IX遺伝子周辺のヒストンの修飾状態をクロマチン免疫沈降法で解析した。脳だけでヒストンが活性化されている状態にあることが分かる。
- B: ゲルシフト法を用い、GnT-IX遺伝子近傍のDNA領域に結合する転写因子を解析した。結合する可能性のある転写因子に対するさまざまな抗体(αCTCFなど)を反応液に添加したところ、転写因子とDNAが結合している場合には、抗体がその結合を妨げ、バンドが消失する様子が分かる。これにより結合する転写因子としてCTCFとNeuroD1を同定した。
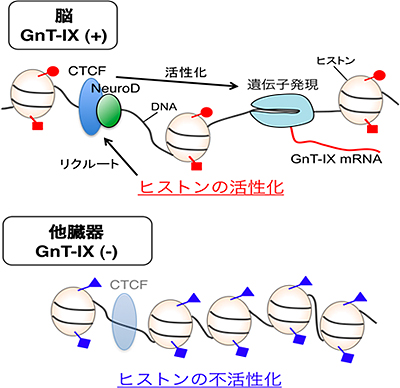
図3 脳に特異的なGnT-IX遺伝子の発現の仕組み
脳ではGnT-IX遺伝子周辺のヒストンが活性化している(赤の○、赤の□はアセチル化などの活性化の指標となる修飾を、青の△、青の□はK27トリメチル化などの不活性化の指標となる修飾を表す)。その結果、CTCFとNeuroDという2つの転写因子が呼び込まれる。結合した2つの転写因子は、GnT-IX遺伝子の発現を活性化する。GnT-IX遺伝子が発現しない他の臓器では、遺伝子周辺のヒストンが不活性化の状態にあり、2つの転写因子の結合も弱く、遺伝子発現が抑制されている。