2011年10月25日
独立行政法人 理化学研究所
ついに植物ホルモン「オーキシン」生合成の主経路を解明
-農作物やバイオマスなどの増収研究に向けて大きな一歩-
ポイント
- 植物はアミノ酸のトリプトファンから2種類の酵素の働きでオーキシンを合成
- 化学的に不安定な極微量のオーキシン生合成中間物質の分析法を確立
- オーキシンによる植物の基本的な成長制御機構の解明に貢献
要旨
独立行政法人理化学研究所(野依良治理事長)は、植物ホルモン※1の1種「オーキシン※2」の生合成の主経路の解明に成功しました。これは、理研植物科学研究センター(篠崎一雄センター長)生長制御研究グループ(神谷勇治グループディレクター)の笠原博幸上級研究員らを中心とした国際共同研究グループ※3による成果です。
オーキシンは植物の形態形成で中心的な役割を果たす成長制御物質です。オーキシンの研究は19世紀後半にダーウィン親子が行った植物の光屈性試験に始まり、1930年代中頃にインドール-3-酢酸(IAA)がオーキシンとして最初に同定されました。植物では1946年にIAAが確認され、その後60年以上にわたりIAAを中心にオーキシンの研究が行われてきましたが、植物がどのようにIAAを合成しているのかは謎のままでした。なぜならIAAの欠損変異体の単離やその生合成中間物質の分析などが困難だったからです。
研究グループは液体クロマトグラフィー・エレクトロスプレーイオン化・タンデム型質量分析装置※4(LC-ESI-MS/MS)という質量分析計※5を使って、植物にごく微量にしか存在せず化学的にも不安定なIAA生合成途中に生成される中間物質(生合成中間物質)を調べるため、モデル実験植物であるシロイヌナズナ※6を使って分析しました。その結果、これまで異なるIAA生合成経路に存在すると考えられていたトリプトファンアミノ基転移酵素(TAA1)とフラビンモノオキシゲナーゼ※7(YUCCA)が、実際は同じ経路に存在する酵素であることを突き止めました。また、YUCCAがIAA生合成中間物質であるインドール-3-ピルビン酸からIAAを生成することを証明しました。これまでの知見とあわせると、植物はアミノ酸であるトリプトファンからTAA1とYUCCAの2種類の酵素の働きでIAAを合成することが明らかになりました。この成果により、IAAによる成長制御機構をさらに詳しく調べることが可能になります。また、人工的に合成されたオーキシンは除草剤や果実成長促進剤等として広く用いられており、農作物やバイオマスなどの増収研究に貢献すると期待できます。本研究成果は米国科学アカデミー紀要『Proceedings of the National Academy of Sciences of the United States of America:PNAS』のオンライン版(10月24日付け:日本時間10月25日)に掲載されます。なお、米国カリフォルニア大サンディエゴ校との共同研究で分子遺伝学の観点から行ったIAA生合成の研究論文(英語)も同号に掲載されます。
背景
オーキシンの1つであるインドール-3-酢酸(IAA)は、植物内で活発に細胞間を極性輸送※8され、オーキシン受容体タンパク質に働き掛けることで、オーキシン早期応答遺伝子の転写活性を促進してオーキシン応答を引き起こします。こうしたIAAの作用機構は明らかにされてきたものの、植物がIAAをどのように合成しているのかという根本的な問題はこれまで解明されていませんでした。IAAが複数の経路から合成されているため、その欠損変異体を単離するのが難しいと考えられていたことが原因です。また、植物にごく微量に存在するIAA生合成中間物質は化学的に不安定なため、分析することが難しかったことも原因に挙げられます。植物の基本的な成長制御の仕組みを理解するうえでオーキシン生合成経路の解明は重要であり、長年にわたる生物学の中心的課題でした。
シロイヌナズナにはアミノ酸であるトリプトファンから始まる4つのIAA生合成経路が存在する可能性があります。研究グループは2009年に、このうちの1つの経路が植物に共通したIAA生合成経路ではなく、アブラナ科固有の二次代謝経路から分岐する特殊な経路であることを示しました(図1 点線内、2009年3月プレスリリース)。一方、IAA生合成酵素は、これまでにいくつも同定されており、なかでも植物全般的に重要な役割を持つTAA1とYUCCAは、別々のIAA生合成経路に存在すると考えられていました(図1左)。しかし、これらの酵素をコードする遺伝子を欠損した変異体が類似した表現型を示すことから、研究グループは、この2つの酵素が同じIAA生合成経路に存在するのではないかと予想しました。
研究手法と成果
YUCCAはIAA生合成の量を制御する鍵となる酵素であり、その遺伝子を過剰発現したシロイヌナズナはIAA過剰生産の表現型を示します。そこで、研究グループはTAA1がYUCCAと同じ経路に存在するならば、2つの遺伝子を同時に発現(共発現)させることによりYUCCA単独の場合よりも過剰にIAAを生産するのではないかと考えました。これら2つの遺伝子の共発現体を作成すると、予想通りTAA1がYUCCAによるIAAの生合成を顕著に増加させることが明らかになりました(図2)。これにより、TAA1とYUCCAが同一経路に存在する可能性が示唆されました。
次に、TAA1遺伝子やYUCCA遺伝子の過剰発現体や欠損変異体のIAA生合成中間体をLC-ESI-MS/MSを使って分析すると、TAA1は報告された通りアミノ酸のトリプトファンからインドール-3-ピルビン酸を合成する酵素ですが、YUCCAはN-ヒドロキシトリプタミンの合成ではなく、インドール-3-ピルビン酸からIAAを合成する酵素である可能性が示唆されました。そこで、大腸菌で調製したYUCCAタンパク質を使って酵素活性試験を行うと、インドール-3-ピルビン酸からIAAを生成することが確認されました。これにより、IAAはトリプトファンからTAA1とYUCCAの2つの酵素の働きにより同一経路上で合成されていることが明らかになりました(図1右)。
今後の期待
LC-ESI-MS/MSによるオーキシン生合成中間体の分析とTAA1遺伝子およびYUCCA遺伝子発現の解析により、植物がいつ、どこで、どの程度のIAAを合成しているのか、IAAによる形態形成や環境応答機構の解明が進むと考えられます。また、人工的に合成されたオーキシンは除草剤や着果・果実成長促進剤、発根促進剤として農業分野で極めて重要であることから、インドール-3-ピルビン酸経路が新たな農薬開発のターゲットになると考えられます。さらに、TAA1遺伝子やYUCCA遺伝子を制御することにより植物でのIAA内生量をコントロールして、人工的に合成されるオーキシンを使用することなく、農作物、綿花などの衣料原料、樹木バイオマスなどを増産する新たな研究の道が拓かれると期待できます。
原論文情報
- Kiyoshi Mashiguchi, Keita Tanaka, Tatsuya Sakai, Satoko Sugawara, Hiroshi Kawaide, Masahiro Natsume, Atsushi Hanada, Takashi Yaeno, Ken Shirasu, Hong Yao, Paula McSteen, Yunde Zhao, Ken-ichiro Hayashi, Yuji Kamiya and Hiroyuki Kasahara “The main auxin biosynthesis pathway in Arabidopsis”. Proceedings of the National Academy of Sciences of the United States of America, 2011, doi:10.1073/pnas.1108434108
発表者
理化学研究所
植物科学研究センター 生長制御研究グループ
グループディレクター 神谷 勇治(かみや ゆうじ)
上級研究員 笠原 博幸(かさはら ひろゆき)
Tel: 045-503-9660 / Fax: 045-503-9662
お問い合わせ先
横浜研究推進部 企画課Tel: 045-503-9117 / Fax: 045-503-9113
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.植物ホルモン
植物の成長を制御する化学物質の総称。一般的に植物ホルモンは、植物でごくわずかしか作られない。これまでに、オーキシン、ジベレリン、サイトカイニン、エチレン、ジャスモン酸、アブシジン酸、ブラシノステロイド、ストリゴラクトン、サリチル酸、CLEペプチドなどが発見されている。 - 2.オーキシン
植物の成長や形態形成などで中心的な役割を果たす植物ホルモン。サイトカイニンと共に植物細胞分裂や細胞分化を制御することや、光や重力による植物の屈曲に関与することで有名。オーキシンは、植物内で活発に細胞間を 極性輸送※8されオーキシン受容体TIR1タンパク質と結合することで、転写抑制因子Aux/IAAタンパク質の分解を促進しAux/IAAタンパク質によって転写速度を低く抑えられていた転写因子ARFタンパク質を活性化、オーキシン早期応答遺伝子の転写活性を促進してオーキシン応答を引き起こす。 - 3.国際共同研究グループ
理研植物科学研究センター生長制御研究グループの増口潔特別研究員、田中慧太ジュニアリサーチアソシエートと、新潟大学の酒井達也准教授、岡山理科大学の林謙一郎准教授、理研植物科学研究センター植物免疫研究グループの白須賢グループディレクター、八丈野孝研究員、東京農工大学大学院連合農学研究科の夏目雅裕教授、川出洋准教授、米国カリフォルニア大学サンディエゴ校のYunde Zhao准教授、米国ミズーリ大学のPaula McSteen准教授らで構成。 - 4.液体クロマトグラフィー・エレクトロスプレーイオン化・タンデム型質量分析装置(LC-ESI-MS/MS)
液体クロマトグラフィー(LC)とは、化合物を分離する技術の1つ。エレクトロスプレーイオン化(ESI)とは、溶液として流れ出てくる個々の化学成分を穏やかな条件でイオン化する方法。タンデム型質量分析装置(MS/MS)とは、目的の化合物に由来するイオンをさらに分解して得られるイオン(フラグメントイオン)から物質を同定・定量する装置。LC-ESI-MS/MSはこれらを組み合わせた質量分析装置で、オーキシンの生合成中間体のようなごく微量で不安定な物質の分析において威力を発揮する。 - 5.質量分析計
物質の正確な分子量を測定する機器。試料をイオン化し、化合物の質量電荷比(質量を電荷数で割った値)から物質を同定・定量する。高感度で物質を検出できるため、植物ホルモンのような微量物質の分析に有用。 - 6.シロイヌナズナ
学名は Arabidopsis thaliana (L.) Heynh. アブラナ科シロイヌナズナ属の1年草。モデル実験植物として植物で初めてゲノム解読が行われ、全遺伝子数は約30,000個。 - 7.フラビンモノオキシゲナーゼ(YUCCA)
フラビンとNADPH/NADHを補酵素とする酸化還元酵素。さまざまな生物に存在する。YUCCAはこの一種で、これまで植物のオーキシン生合成の中でトリプタミンを酸化して N-ヒドロキシトリプタミンを生成する酵素と考えられていた。本研究によりインドール-3-ピルビン酸からIAAを生成する酵素であることが明らかにされた。 - 8.極性輸送
方向性のある物質輸送。植物のオーキシン極性輸送においては、オーキシンを細胞から排出するPINタンパク質が特に重要であることが示されている。
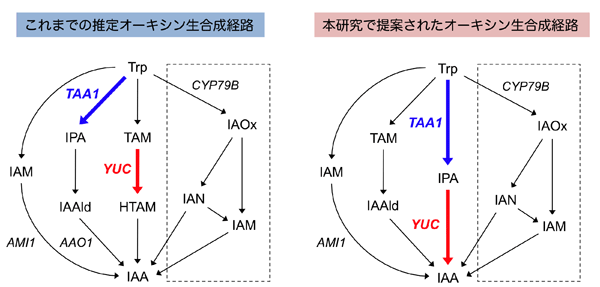
図1.植物のIAA生合成経路
左図:今までシロイヌナズナのインドール-3-酢酸(IAA)生合成経路において、TAA1とYUCCA(YUC)は別々の経路に含まれると予想されていた。
右図:本研究において、この2つの酵素が同じIAA生合成経路に存在することが示された。
点線内はシロイヌナズナに存在するアブラナ科固有の二次代謝経路から分岐したIAA 生合成経路。斜体はシロイヌナズナで遺伝子が単離されているIAA生合成酵素。インドール-3-アセトアルデヒド(IAAld)は本研究で提案された主経路には含まれない。
Trp:トリプトファン、IAM:インドール-3-アセトアミド、IPA:インドール-3-ピルビン酸、TAM:トリプタミン、HTAM:N-ヒドロキシトリプタミン、IAOx:インドール-3-アセトアルドキシム、IAN:インドール-3-アセトニトリル、AMI1:アミダーゼ1、AAO1:アルデヒドオキシダーゼ1。

図2.2種類のIAA生合成酵素(TAA1とYUCCA)の遺伝子を同時に発現させたシロイヌナズナ
TAA1ox YUC6ox:同じ経路に存在するTAA1遺伝子とYUCCA遺伝子を共発現させたシロイヌナズナ。側根が顕著に成長している。
TAA1ox、YUC6ox:TAA1遺伝子とYUCCA遺伝子をそれぞれ単独で過剰発現させたシロイヌナズナ。
pER8:これらの遺伝子を過剰発現していない植物体。