ポイント
- ストリゴラクトンは乾燥ストレスと塩ストレスへの耐性を増大する制御因子
- ストリゴラクトンは気孔の発達と機能を制御し乾燥ストレスに適応
- 植物は環境ストレスに適応するために複合的なホルモン応答経路を作り出す
要旨
理化学研究所(理研、野依良治理事長)は、実験モデル植物のシロイヌナズナを使い、植物ホルモン「ストリゴラクトン(SL)[1]」が、乾燥と高塩濃度ストレスへの耐性を増大させる制御因子であることを明らかにしました。また、SL、アブシジン酸[2]、サイトカイニン[3]による複合的なホルモン応答経路を通じて、ストレス応答遺伝子の発現と気孔の閉鎖により環境ストレスに適応する新たなメカニズムがあることが分かりました。これは、理研環境資源科学センター(篠崎一雄センター長)発現調節研究ユニットのチャン・ソン・ファン・ラム ユニットリーダー、ハ・チエン・バン国際プログラム・アソシエイト、西山りゑ研究員、機能開発研究グループの篠崎一雄グループディレクター、植物ゲノム発現研究チームの関 原明チームリーダーと、メキシコのシンベスタフ・ランヘビオのルイス・エレラ・エストゥレヤ教授らの共同研究グループによる成果です。
乾燥や高塩濃度などの環境ストレスは、植物の成長や生産性に悪影響を及ぼします。これに適応するため、さまざまな植物ホルモンがストレス応答経路で協調して働くことが知られていますが、ストレス応答におけるSLの機能は分かっていませんでした。
共同研究グループは、シロイヌナズナのSLの合成を阻害した低SL変異体と、SLのシグナル伝達を阻害したSLシグナル変異体を用いて、乾燥ストレスと塩ストレスの耐性テストを行いました。その結果、乾燥と高塩濃度のストレスに弱くなることを見いだし、SLがこれらのストレス耐性を増大させる制御因子であることを証明しました。逆に、野生型のシロイヌナズナにSLを与えるとストレス耐性が向上しました。ストレスに対するSLの効果は根ではなく地上部で発揮され、気孔の発達とアブシジン酸感受性に関係していることが知られています。SLを介したストレス応答経路は、ストレス応答遺伝子、アブシジン酸応答遺伝子、サイトカイニン代謝関連遺伝子の発現を制御することにより、気孔を閉鎖して乾燥による水分の蒸散を抑えると考えられます。これらの結果から、植物は環境ストレスに適応するためにSL、アブシジン酸、サイトカイニンによる複合的なホルモン応答経路を作り出したことが示唆されました。今後、SL量を増やしたりSL応答経路を強化したりすることで、環境ストレスによる作物の生産性低下を軽減する新たな育種方法の開発につながると期待できます。
本研究成果は、米国科学アカデミー紀要『Proceedings of the National Academy of Sciences of USA』(2014年1月14日付け)に掲載されました。
背景
乾燥や高塩濃度などの環境ストレスは、植物の成長や生産性に悪影響を及ぼします。これに対応するため、アブシジン酸やサイトカイニン、ストリゴラクトン(SL)など、さまざまな植物ホルモンがストレス応答経路で協調して働くことが知られています。
SLはカロテノイド由来の化合物の1種で、発芽の促進、植物やカビの寄生の促進、腋芽(葉の付け根にできる芽)の成長阻害による枝分かれの抑制に働くことが報告されています。モデル実験植物のシロイヌナズナとイネで、SLの合成酵素遺伝子とシグナル伝達遺伝子は同定されていますが、ストレス応答におけるSLの機能については分かっていませんでした。
研究手法と成果
共同研究グループは、シロイヌナズナのSLの合成を阻害した低SL変異体と、SLのシグナル伝達を阻害したSLシグナル変異体を用いて、乾燥ストレスと塩ストレスの耐性テストを行いました(図1)。その結果、低SL変異体とSLシグナル変異体は乾燥と塩ストレスに感受性を示し(弱くなる)ました。これは、SL量の減少やSLシグナル伝達の阻害がストレス応答に影響していることを示しています。さらに、この低SL変異体のストレス耐性の低下は、SLを与えることで元に戻りました(図2)。また、野生型のシロイヌナズナにSLを与えるとストレス耐性が向上しました(図2)。これらの結果は、SLが乾燥と塩のストレス耐性を増大させる制御因子であることを裏づけています。
また、両変異体では、野生型に比べて乾燥による水分の蒸散が多いことから、SL量の減少とSLシグナル伝達の阻害によって蒸散率が上昇し、乾燥ストレス耐性が弱くなったことが分かりました(図3A)。アブシジン酸によって制御される気孔閉鎖や気孔密度が乾燥ストレスによる水分の蒸散に影響を及ぼすことはすでに知られています。そこで、アブシジン酸と気孔の関係を調べたところ、両変異体では、野生型に比べてアブシジン酸による気孔の閉鎖が起こりにくくなり、気孔密度が高くなっていることが分かりました(図3B、C)。両変異体では、アブシジン酸による気孔閉鎖の阻害と気孔の増加により、乾燥による水分の蒸発量が増加し、乾燥耐性が低くなったと考えられます。
さらに共同研究グループは、SLを介した乾燥ストレス応答の分子メカニズムを解明するため、通常および乾燥条件下で育成したSLシグナル変異体と野生型の葉の転写物解析を行いました。その結果、SLを介した分子メカニズムでは、気孔の機能や光合成、フラボノイド[4]化合物の生合成、サイトカイニン代謝などを制御する多くの遺伝子の発現を調節することによって、ストレス適応をコントロールしていることが明らかになりました。
今後の期待
今回の成果は、環境ストレスに適応するため植物が複数のホルモン応答経路を協調して発達させてきたことを示唆しています。ホルモン間の相互作用がどこでどのように起きるかを突き止めることは、学術的に非常に価値のある研究です。また応用研究の観点からも、本研究結果はSL処理により植物のストレス耐性を増強できることを示しており、環境ストレスに対抗するための新たな道を切り開くことに成功しました。SLを利用したストレス耐性植物の開発には、SLの大規模生産コストを抑える必要があり、今後、産業的視点での研究が鍵になります。遺伝子工学によるSL生合成やSLシグナルの遺伝子操作は、乾燥や高塩濃度のストレス耐性作物の開発のために有望なアプローチになると考えられます。
原論文情報
- Chien Van Ha, Marco A. Leyva-González, Yuriko Osakabe, Uyen Thi Tran, Rie Nishiyama, Yasuko Watanabe, Maho Tanaka, Motoaki Seki, Shinjiro Yamaguchi, Nguyen Van Dong, Kazuko Yamaguchi-Shinozaki, Kazuo Shinozaki, Luis Herrera-Estrella, and Lam-Son Phan Tran. "Positive regulatory role of strigolactone in plant responses to drought and salt stress", Proceeding of the National Academy of Sciences,2013,doi: 10.1073/pnas.1322135111.
発表者
理化学研究所
環境資源科学研究センター 発現調節研究ユニット
研究員 西山 りゑ(にしやま りえ)
国際プログラム・アソシエイト ハ・チエン・バン
ユニットリーダー TRAN Son Phan Lam(チャン・ソン・ファン・ラム)
お問い合わせ先
環境資源科学研究推進室 広報担当者
Tel: 048-467-9449 / Fax: 048-465-8048
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.ストリゴラクトン(SL)
植物体内で合成され発芽や成長などの生理過程を調節する化合物を植物ホルモンと呼ぶ。ストリゴラクトンはその1種で発芽の促進や腋芽の成長阻害による枝分かれの抑制などに関与する - 2.アブシジン酸
種子の成熟や休眠、落葉などに関与する植物ホルモン。その他、気孔の閉鎖を誘導し水分の蒸散を抑える、細胞の水分状態を保つ働きを持つ防御物質の蓄積を促進するなどの作用があり、乾燥や塩ストレスに対するポジティブレギュレーターであることが分かっている。 - 3.サイトカイニン
茎や葉の成長を促進し、根の成長を阻害する性質を持つ植物ホルモン。細胞分裂や老化、側芽の成長などにも関与している。2011年に理研の研究グループにより、サイトカイニンはストレス応答のネガティブレギュレーターとして働くこと、サイトカイニンとアブシジン酸の相互作用がストレス応答を制御することが報告されている。 - 4.フラボノイド
天然に存在する植物二次代謝化合物で、アントシアニンやカテキンなどのポリフェノール類の総称。ストレスに応答して合成される防御物質の1種と考えられている。

図1 低SL変異体とSLシグナル変異体の乾燥・塩ストレス処理
- A: 環境ストレスにさらさずに土壌で育てたシロイヌナズナ。左上は低SL変異体、左下はSLシグナル変異体、右は野生型。
- B: 乾燥耐性テストの結果。Aの水遣りを止めて乾燥させると、野生型は生き延びるが、両変異体のほとんどは枯死する。
- C: 塩耐性テストの結果。Aに塩水(200mM NaCl)を与えると、野生型は生き延びるが、両変異体のほとんどは枯死する。
- D: 乾燥・塩ストレス処理での生存率。野生型に比べ、両変異体は低い生存率を示す。
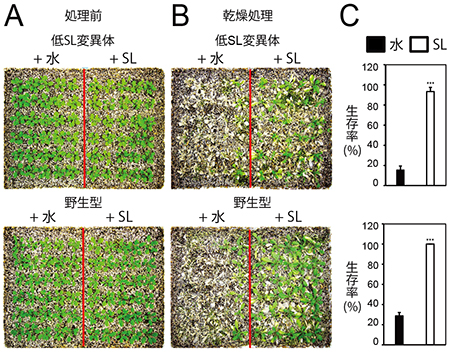
図2 低SL変異体と野生型にSLを与えた結果
- A: 環境ストレスにさらさずに土壌で育てた低SL変異体(上)と野生型(下)のシロイヌナズナ。
- B: SLを与えた結果。Aの水遣りを止めて乾燥させると同時に、5μMのSL(右)あるいは水(左)を葉にスプレーする。その結果、低SL変異体、野生型ともに、SL処理した個体だけが生き延びる。
- C: SLを与えた際の生存率。SLを与えなかった個体(水を与えただけの個体)に比べ、SLを与えた個体は高い生存率を示す。
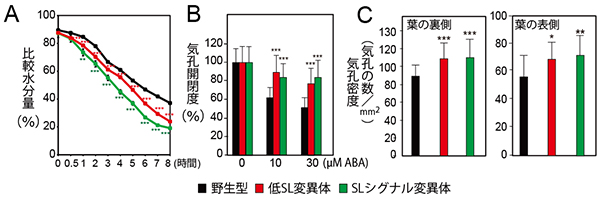
図3 気孔の発達とアブシジン酸感受性の増加
- A: 低SL変異体とSLシグナル変異体、野生型の乾燥ストレスによる水分量の比較。育成培地から取出し、乾燥させながら植物体の重量を測定することで水分損失率の変化を見ることができる。両変異体は野生型に比べて、乾燥による水分の蒸散が多いことが分かる。
- B: アブシジン酸に対する気孔開閉度の比較。野生型に比べて両変異体は、アブシジン酸による気孔の閉鎖が起こりにくくなっている。
- C: 気孔密度の比較。野生型に比べて両変異体は、葉の両面で気孔密度が高くなっている。