2014年6月9日
理化学研究所
抗体を産生する免疫応答に重要なT細胞の動きを制御する仕組みを解明
-脂質メディエーターの受容体(S1PR2)によるT細胞の局在制御-
ポイント
- S1PR2とCXCR5の働きによって濾胞性ヘルパーT細胞が適切な場所に局在する
- この局在が無くなると、胚中心反応が減弱する
- より良い抗体の長期的な産生を目的としたワクチン開発や改良への応用に期待
要旨
理化学研究所(理研、野依良治理事長)は、良質な抗体を長期に産生する免疫反応「胚中心反応[1]」に必要なT細胞[2]の動きが、リンパ節や脾臓などの免疫組織の中でどのように制御されているのかを明らかにしました。これは、理研統合生命医科学研究センター(小安重夫センター長代行)組織動態研究チームの岡田峰陽チームリーダー、森山彩野基礎科学特別研究員を中心とする共同研究チームの成果です。
私たちの体には、生体を脅かす細菌やウイルスなどの外敵(抗原)に対抗するため、免疫応答という防御機能が備わっています。体内に侵入した抗原を排除するためにB細胞[3]が抗体を産生する反応もその1つです。さらに、B細胞はその局在領域である濾胞[4]の中に作られる胚中心と呼ばれる場所で自らが作り出す抗体の性能を高め、抗原の再侵入に対して備える胚中心反応を起こします。胚中心反応には、T細胞の一種である「濾胞性ヘルパーT細胞(TFH細胞)[5]」の助けが必要です。これまで、TFH細胞が移動して濾胞の中で行われるB細胞の胚中心反応に加わるためには、TFH細胞表面に発現しているケモカイン受容体[6]「CXCR5」が重要であることが知られていました。しかし、CXCR5を欠損した場合でもTFH細胞が胚中心から無くなりません。このため、共同研究チームはケモカイン受容体以外にもTFH細胞の移動を制御する因子が存在すると考え、TFH細胞がどのように胚中心に局在するのか、また、胚中心TFH細胞が無くなった時にB細胞の反応がどう変化するのか調べました。
共同研究チームは、脂質から作られる生理活性因子「脂質メディエーター」の1つであるスフィンゴシン1リン酸(S1P)[7]の受容体「S1PR2」に着目しました。遺伝子改変マウスを用いた実験の結果、S1PR2がCXCR5と協力して胚中心TFH細胞の移動を制御していることを発見しました。また、胚中心TFH細胞の存在が、胚中心の形成や維持に重要であることも示されました。この成果を応用することで、将来的に、免疫細胞の組織内での動きの制御が可能になれば、より良い抗体の産生を目的としたワクチン療法の新規開発や改良が図れると期待できます。
本研究成果は、科学技術振興機構 戦略的創造研究推進事業 個人型研究(さきがけ)研究領域「炎症の慢性化機構の解明と制御」(研究総括:高津 聖志 富山県薬事研究所 所長)における研究課題「免疫・炎症研究におけるオプトジェネティクスの創生」(研究者:岡田峰陽)の一環として行われ、米国の科学雑誌『The Journal of Experimental Medicine』オンライン版(6月9日付け:日本時間6月9日)に掲載されます。
背景
私たちは、常に細菌やウイルスなどの外敵(抗原)にさらされています。これら抗原に対抗するため、私たちの体には免疫応答という生体防御機能が備わっています。免疫応答の中でも重要なものの1つが、体内に侵入した抗原を排除するためにB細胞が抗体を産生する反応です。さらに、B細胞はその局在領域である濾胞の中に作られる胚中心と呼ばれる場所で、自らが作り出す抗体の性能を高め、抗原の再侵入時に素早く対応できるように備える「胚中心反応」を起こします。近年注目されている、ヒト免疫不全ウイルス(HIV)やインフルエンザウイルスといった変異性の高いウイルスを中和することのできる抗体も、胚中心で作られると考えられています。
胚中心反応には、T細胞の一種である濾胞性ヘルパーT細胞(TFH細胞)の助けが必要です。普段、T細胞は免疫組織の中でT細胞領域に局在しており、B細胞が局在する濾胞とは別の場所に存在します。しかし抗原が侵入すると、活性化したT細胞の一部がTFH細胞となり、B細胞がいる濾胞へ移動します(図1)。この移動には、TFH細胞表面に発現しているケモカイン受容体「CXCR5」が重要であることが知られていました。また、TFH細胞の一部は胚中心の中にも見られ、胚中心B細胞の反応を助けていると考えられていました。しかし、CXCR5が欠損していても、TFH細胞は胚中心から無くならないため、TFH細胞がどうやって胚中心の中に局在しているのか、また、胚中心TFH細胞が無くなった時にB細胞の反応がどのように変化するかが議論になっていました。しかし、この問題はいまだに解明されていません。
共同研究チームは、これまでに、生理活性因子「脂質メディエーター」の1つであるスフィンゴシン1リン酸(S1P)の受容体「S1PR2」が、他のT細胞に比べてTFH細胞で強く発現していることを発見していました。今回、共同研究チームは、S1PR2の発現とその働きを詳細に解析することで、S1PR2が胚中心TFH細胞の局在を制御するメカニズムと、胚中心TFH細胞が胚中心反応で果たす役割を明らかにしようと試みました。
研究手法と成果
共同研究チームは、まず、TFH細胞でのS1PR2の発現を可視化するために、S1PR2をコードする遺伝子「S1pr2」を発現した細胞が蛍光を発する遺伝子改変マウスを作製しました。この遺伝子改変マウスを用いて、TFH細胞におけるS1pr2遺伝子の発現をフローサイトメトリー法[8]で調べたところ、発現は一様ではなく一部のTFH細胞だけがより強くS1pr2遺伝子を発現していました。また、リンパ節の組織切片上でS1pr2遺伝子を強く発現しているT細胞を観察したところ、そのほとんどが胚中心の中に観察されました。これらの結果から、S1PR2は胚中心TFH細胞に強く発現することが示唆されました。
さらに、S1PR2がTFH細胞の胚中心局在を制御するかどうかを明らかにするため、リンパ節の組織切片上で野生型マウス由来のT細胞とS1pr2遺伝子欠損マウス由来のT細胞の局在を観察し、比較しました。その結果、S1pr2遺伝子欠損マウスのT細胞は、野生型マウスのT細胞に比べて胚中心局在が半分程度まで減少していました。この現象をさらに詳細に解析するため、二光子励起レーザー顕微鏡を用いて胚中心とその周辺のライブイメージング[9]を行ったところ、野生型マウスの胚中心TFH細胞は胚中心内に留まろうとするのに対して、S1pr2遺伝子欠損マウスのTFH細胞は、胚中心内に留まろうとする動きが全く観察されませんでした(図2)。これらのことから、S1PR2は胚中心TFH細胞が胚中心内に留まるために必要な因子であることが示唆されました(図1)。
共同研究チームは、胚中心TFH細胞の局在制御メカニズムと胚中心反応における役割をさらに詳細に調べるために、CXCR5を欠損したマウスと、CXCR5とS1PR2の両方を欠損したマウスを用い、実験を行いました。過去の報告からT細胞には、胚中心を形成し維持する能力があることが分かっています。実験の結果、CXCR5だけを欠損したT細胞には、野生型T細胞と比べると弱いながらも胚中心を形成し維持していく能力があることが確認されました。一方、CXCR5とS1PR2を両方欠損したT細胞は、胚中心を形成し維持することがほとんどできないことが分かりました。さらに野生型T細胞の共存下で胚中心を形成させ、これらのT細胞の局在を調べたところ、CXCR5だけを欠損したT細胞は胚中心の中に観察されましたが、CXCR5とS1PR2の両方を欠損したT細胞は、胚中心の中にほとんど観察されませんでした。これらの結果から、S1PR2はCXCR5とともに、T細胞の胚中心への局在を制御していること、また、胚中心TFH細胞の存在が胚中心の形成や維持に必要であることが示唆されました。
今後の期待
共同研究チームは、胚中心TFH細胞の局在がS1PR2とCXCR5により制御され、これらによる胚中心TFH細胞の適切な局在が、胚中心反応に重要であることを明らかにしました。今回の成果を応用し、将来的に胚中心TFH細胞の局在を制御できれば、より良い抗体の長期的な産生を目的としたワクチン療法の新規開発や改良が図れると期待できます。
原論文情報
- Saya Moriyama, Noriko Takahashi, Jesse A. Green, Shohei Hori, Masato Kubo, Jason G. Cyster, and Takaharu Okada. “Sphingosine-1-phosphate receptor 2 is critical for follicular helper T cell retention in germinal centers”, The Journal of Experimental Medicine, 2014, doi: 10.1084/jem.20131666
発表者
理化学研究所
統合生命医科学研究センター 組織動態研究チーム
チームリーダー 岡田 峰陽(おかだ たかはる)
お問い合わせ先
統合生命医科学研究推進室
Tel: 045-503-9117 / Fax: 045-503-9113
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.胚中心反応
胚中心は、主にリンパ節や脾臓などの免疫組織において、免疫応答中に濾胞の中に作られる反応場。ここでは、免疫応答中の増殖B細胞が、自己の抗体遺伝子に変異を入れることで、抗原に対する親和性を向上させていく。これを胚中心反応と呼ぶ。この反応を介して生まれる抗体産生細胞は、その後骨髄に移動して長期間生存することができる。外敵を駆逐するために非常に重要な反応である一方、遺伝子変異が高頻度で起こる反応のため、制御に異常が起こるとリンパ腫や自己免疫疾患などの病状を引き起こすと考えられている。 - 2.T細胞
免疫細胞の一種。直接他の細胞と接触し、また、サイトカインと呼ばれる液性因子を分泌することで、B細胞や他の免疫細胞の細胞分化や機能を調節する。分泌するサイトカインの種類や局在場所の違いによって機能が異なる。 - 3.B細胞
免疫細胞の一種。B細胞抗原受容体と呼ばれるタンパク質を細胞表面に出し、抗原を認識する。体の中には100万種以上の異なった抗原認識能力を持つB細胞が存在し、侵入してくるあらゆる病原体やウイルスに対応することができる。最終的にB細胞は、細菌やウイルスを排除するための抗体(分泌型に変化したB細胞抗原受容体)を分泌する抗体産生細胞に分化する。また抗体産生細胞へと分化する前に、胚中心反応を起こして、抗原に対してより親和性の高い抗体を作る。 - 4.濾胞
リンパ節やパイエル板といったリンパ組織で見られる、B細胞が集まった構造のこと。リンパ組織では、B細胞の存在する領域とT細胞の存在する領域ははっきりと分かれており、B細胞は濾胞内に存在し、T細胞は濾胞を取り囲むように存在する。 - 5.濾胞性ヘルパーT細胞(TFH細胞)
T細胞の中でも、T細胞の集合場所(T細胞領域)から濾胞へ移動してB細胞の働きを補助できるように変化した細胞。細胞表面に存在する分子を介した相互作用や、サイトカインと呼ばれる液性因子を分泌してB細胞の働きを調節する。 - 6.ケモカイン受容体
ケモカインは細胞の移動を制御する液性因子。ケモカイン受容体にケモカインが結合すると、その刺激が細胞内部へ伝わり、細胞の移動能が変化する。 - 7.スフィンゴシン1リン酸(S1P)
生体膜の構成成分である脂質の1つから作られる液性因子。S1PR1からS1PR5まで5種類の受容体が存在する。 - 8.フローサイトメトリー法
単離した大量の細胞を1列に並ぶように高速で流しながら、その1つひとつの大きさや分子発現状態について解析する方法。 - 9.二光子励起レーザー顕微鏡を用いたライブイメージング
レーザー蛍光顕微鏡の一種である二光子励起レーザー顕微鏡を用いて、生きている細胞や組織の観察をすること。二光子励起レーザー顕微鏡を使用することで、生体への光による毒性を軽減しつつ、高解像度で3次元画像を取得できる。
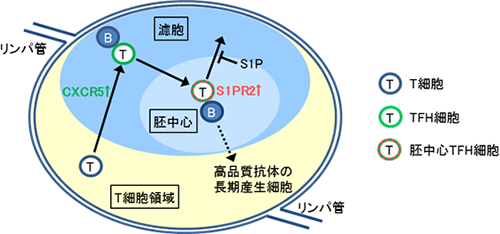
図1 リンパ節内におけるT細胞およびTFH細胞の動き
普段、T細胞は免疫組織の中でT細胞領域に局在している。しかし抗原が侵入すると、活性化したT細胞の一部がTFH細胞となり、B細胞がいる濾胞へ移動する。この際、TFH細胞表面に発現しているケモカイン受容体「CXCR5」が重要であることが知られていた。今回の研究により、胚中心TFH細胞が胚中心内に留まるためにS1PR2が必要な因子であることが示唆された。
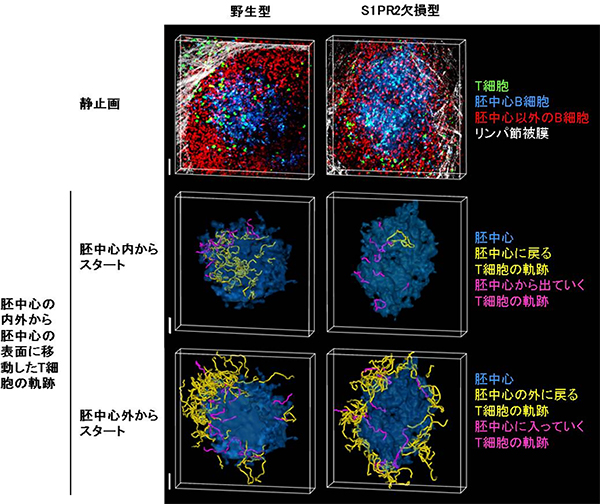
図2 胚中心ライブイメージングの静止画及びT細胞の軌跡
- 上段:ある瞬間における、二光子励起レーザー顕微鏡を用いた胚中心とその周辺のライブイメージング。
- 中段:胚中心内から胚中心表面に移動した野生型T細胞のほとんどは胚中心に戻り留まろうとするが、S1PR2欠損型T細胞はその傾向が見られない。
- 下段:胚中心外から胚中心表面に移動したT細胞は、遺伝子型に関わらずほとんどが胚中心外へ戻る。
スケールバーは50μm。(原論文より改変)