2015年9月30日
理化学研究所
マウスの「父性の目覚め」に重要な脳部位を発見
―オスマウスの子育て意欲は2つの脳部位の活性化状態に表れる―
要旨
理化学研究所(理研)脳科学総合センター親和性社会行動研究チームの黒田公美チームリーダー、恒岡洋右研究員(研究当時)、時田賢一研究員らの研究チーム※は、オスマウスの子育て(養育行動[1])意欲が「cMPOA」と「BSTrh」の2つの脳部位の活性化状態から推定できることを発見しました。
ほ乳類の子は未発達な状態で生まれ栄養源を母乳に頼るため、親による養育が不可欠です。メスマウスは若い時から子の世話をすることが多く、さらに母親になる時には出産時の生理的変化[2]により養育行動が強化されます。一方、交尾未経験のオスマウスは養育せず、子に対して攻撃的ですが、メスとの交尾・同居を経て父親となると、よく養育するようになります(父性の目覚め)。黒田チームリーダーらは2013年に、フェロモンを検出する鋤鼻器(じょびき)[3]の阻害がオスマウスの子への攻撃を抑え、養育を促すことを見いだしています注1)。しかし「父性の目覚め」現象は鋤鼻器が退化している類人猿でも見られることから、さまざまな感覚入力を受けとり子への攻撃や養育行動を制御する、類人猿にも共通する脳領域のメカニズムが重要であると考えられました。
研究チームは今回、オスマウスの子に対する攻撃と養育に必要な中枢の脳領域の同定を試みました。そして、交尾未経験オスの子への攻撃行動には前脳にある分界条床核BST[4]の一部であるBSTrhが重要であり、また父親マウスの養育行動には内側視索前野中央部cMPOA[5]が必要であることを突き止めました。また、cMPOAはBSTrhの働きを抑えること、cMPOAを光遺伝学的手法[6]により活性化すると交尾未経験オスマウスの子への攻撃が減弱すること、メスとの交尾経験後はcMPOAが活性化したことから、父親になる時にはBSTrhに対しcMPOAの活動が優位になることで、子への攻撃をやめ養育する「父性の目覚め」が起こる可能性が示唆されました。さらに、あるオスマウスが子を攻撃するか、養育するかは、cMPOAとBSTrhの2つの脳部位の活性化状態を測定するだけで、95%以上の高精度で推定できることが分かりました。
本研究は、子に対する攻撃と養育という正反対の行動のそれぞれに必要な中枢の脳部位を詳細に同定し、その活性化状態からマウスの行動意欲が読み取れることを示した初めての研究成果です。こういった脳部位の働きを霊長類において調べることで、人間の父子関係の理解とその問題解決に役立つ知識を得ることにつながると考えられます。
本研究成果は、国際科学誌『The EMBO Journal』(9月30日付け:日本時間10月1日)に掲載されます。
注1)2013年3月20日プレスリリース マウスが父性行動を発現する神経機構の一端が明らかに
※研究チーム
理化学研究所 脳科学総合研究センター
親和性社会行動研究チーム
チームリーダー 黒田 公美(くろだ くみ)
研究員 恒岡 洋右(つねおか ようすけ)(研究当時)(東邦大学医学部 助教)
研究員 時田 賢一(ときた けんいち)
神経回路・行動生理学研究チーム
チームリーダー トーマス・マックヒュー(Thomas J McHugh)
背景
ほ乳類の子は、未発達な状態で生まれます。そのため親は、子の生存率を高めるために子育て(養育行動)をします。げっ歯類の養育行動は、子を保温する、身体をなめて清潔に保つ、巣を作る、子を巣に回収する、授乳するといったさまざまな行動からなります。一般に実験室で飼育されるメスマウスは、交尾未経験であっても授乳以外の養育行動を示します。一方で、交尾未経験のオスマウスは子に対して攻撃行動を示します。これは、他のオスの子を排除することにより、授乳中は抑制されているメスの発情を促し、自らの生殖成功率を高めるからであると考えられています。しかし、オスマウスは父親になると攻撃をやめ、自分の子ばかりか他のオスの子に対しても区別なく養育するようになります(図1)。
黒田チームリーダーを含む理研の研究グル―プは2013年に、子の発するフェロモン[7]がオスの子への攻撃に必要であること、また、父親マウスではフェロモン検出を行う鋤鼻器の働きが抑制されることで子への攻撃行動が抑えられていることを見いだしました。しかし「父性の目覚め」現象は鋤鼻器が退化している狭鼻猿類[8]でも見られることから、さまざまな感覚入力を受けて子への攻撃行動と養育行動を制御する、狭鼻猿類にも共通する脳部位のメカニズムが重要であると考えられます。また、2013年の論文では、攻撃と養育の際にどのような中枢の脳部位が活性化するのか、ある程度は分かりましたが、そのうちのどこが機能的に重要なのかはまだ分かりませんでした。
そこで研究チームは今回、子への攻撃および養育時にどのような中枢の脳領域が必要なのかを特定しようと試みました。
研究手法と成果
研究チームはまず、子を攻撃する交尾未経験オスマウスと養育するオスマウスの2つのグループに対して、子と2時間同じケージ内に同居させたときに、脳のどの領域が活性化しているかを、神経細胞の活動の指標であるc-Fosタンパク質[9]を用いて調べました。対照群として、2時間単独でいたオスマウスのグループと比較したところ、攻撃する際にはオスマウスの前脳の分界条床核BSTの一部であるBSTrhという部位で、養育する際には内側視索前野中央部cMPOAという部位で、それぞれc-Fos陽性細胞密度が増加し、有意に活性化していることが分かりました(図2)。また、子を金網で覆い、実際には攻撃や養育ができない状況(間接呈示)にしても、それぞれの行動に対応した活性化が見られました。この結果から、これらの2つの脳部位の活性化は、子に対して起こした行動の結果ではなく、行動を起こそうとする意欲や動機を反映していると考えられました(図2)。
研究チームは、cMPOAとBSTrhの2つの脳部位のc-Fos陽性細胞の密度を使って活性化状態を調べるだけで「あるオスマウスが子を攻撃するのか養育するのか」を推定できるか調べました。統計学的な手法の1つである樹木モデル解析の結果、95%以上の高い精度でどちらの行動をとるかを推定できることが分かりました(図3)。具体的には、どちらの行動をとるのか分からないオスマウスについて、BSTrhに発現するc-Fos陽性細胞の密度を調べたとき、その値が187.8よりも大きい場合には攻撃するマウスであることが、真陽性率[10]100%、偽陽性率[10]0%という非常に確かな精度で推定できました。さらに、BSTrhのc-Fos陽性神経細胞の密度が187.8以下である場合は、cMPOAのc-Fos陽性細胞の密度を調べます。その値が31.9よりも高ければ養育するマウスであると、真陽性率92.6%、偽陽性率2.5%という、かなり高い精度で推定することができました。また、直接呈示と間接呈示の違いは、結果に影響を与えませんでした。このことから、実際に行動したかどうかに関わらず、オスマウスが子に対して攻撃するのか、それとも養育するのか、脳の活性化状態から推測できるということが分かりました。また、BSTrhやcMPOAの活性化は、行動の結果ではなく「行動への意欲」で起きることが分かりました。
次に、cMPOAとBSTrhの神経活動が攻撃行動や養育行動に必要かどうかを確かめるため、それぞれの働きを阻害する薬剤を用いた実験を行いました。交尾未経験のオスマウスのBSTrhの働きを阻害すると、BSTrhの働きを阻害しなかった交尾未経験オスマウスに比べ、子への攻撃行動が有意に減少しました(図4)。これは、BSTrhの働きが攻撃行動を促進することを示唆します。一方で、父親マウスのcMPOAの働きを阻害すると、全く養育しなくなっただけでなく、子を攻撃するようになりました(図4)。
また、cMPOAの働きを阻害するとBSTrhが活性化されたことから、cMPOAはBSTrhを抑制していることが分かりました。つまり、養育行動に関わるcMPOAが活性化されると、攻撃行動に関わるBSTrhの働きが抑えられるような神経回路を形成しているといえます。さらに、この2つの部位の結合様式を解析すると、実際にGABA作動性の抑制性神経[11]がcMPOAからBSTrhへ投射していることが分かりました。
最後に、cMPOAの活性化が攻撃行動へ与える影響を最新の光遺伝学的手法を用いて調べました。研究チームは、ウイルスベクターを利用して、光が当たると神経細胞を活性化するチャネルロドプシン[12]というタンパク質をcMPOAの神経細胞にだけ作らせることにより、交尾未経験のオスマウスのcMPOAを光で活性化しました。すると子に対する攻撃行動が有意に減少したことから、実際にcMPOAの活性化は攻撃行動を抑制することが示されました(図5)。また、父親になるためにはメスと交尾する経験が必要です。そこで、メスと交尾する経験がcMPOAに与える影響を検討するため交尾して2時間後のオスマウスを調べたところ、cMPOAが活性化していました。
以上の結果から、オスマウスがメスとの交尾・同居を経て父親になる際には、まず交尾によってcMPOAの活性が高まり、cMPOAが攻撃行動を制御しているBSTrhの神経活動を抑制することによって攻撃行動を抑え、養育行動を促進している可能性が示されました(図6)。
今後の期待
本研究では、オスマウスの子に対する攻撃と養育という、正反対の行動に重要な中枢の脳部位BSTrhとcMPOAを同定しました。また、cMPOAとBSTrhの活性化状態からマウスの行動意欲が読み取れることを初めて示しました。
養育行動に必須の脳内部位cMPOAは、子が生まれる前の、メスとの交尾・同居という社会的つながりによってオスの脳内で活性化され、子に対する攻撃行動を抑制します。そして子が生まれると、その経験によって父親は子育てを学んでいくと考えられます。
今回明らかになったマウスでの「父性の目覚め」のメカニズムは、すぐに人間に応用することはできません。また、現在の技術では、cMPOAやBSTrhのような微小な脳深部の脳神経核の活動を非侵襲的に測定することは困難です。しかし、cMPOAやBSTrhなどの脳領域は哺乳類の中で進化的に保存されているため、こういった脳部位の働きを霊長類において調べることで、人間の父子関係の理解とその問題解決に役立つ知識を得ることができると期待されます。
原論文情報
- Yousuke Tsuneoka, Kenichi Tokita, Chihiro Yoshihara, Taiju Amano, Gianluca Esposito, Arthur J. Huang, Lily M.Y. Yu, Yuri Odaka, Kazutaka Shinozuka, Thomas J. McHugh, and Kumi O. Kuroda, "Distinct preoptic-BST nuclei dissociate paternal and infanticidal behavior in mice", The EMBO Journal, doi: 10.15252/embj.201591942
発表者
理化学研究所
脳科学総合研究センター 親和性社会行動研究チーム
チームリーダー 黒田 公美 (くろだ くみ)
研究員 恒岡 洋右 (つねおか ようすけ)(研究当時)
研究員 時田 賢一 (ときた けんいち)
 黒田 公美
黒田 公美
 恒岡 洋右
恒岡 洋右
 時田 賢一
時田 賢一
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.養育行動
子の生存の可能性を高めるような親の行動であり、ほ乳類をはじめとして、鳥類、は虫類、両生類、魚類、さらに無脊椎動物にも広く見られる行動である。ほ乳類ではほ乳の必要性から母親による養育は例外なく行われる。 - 2.出産時の生理的変化
妊娠時は胎盤からエストロゲン、プロゲステロンが分泌されるが、分娩により急減する。また、乳腺発達や乳汁分泌を促進するプロラクチンは、妊娠時から分娩後の授乳期にかけて、脳下垂体前葉から分泌される。分娩中・分娩後には子宮収縮作用・射乳作用のあるオキシトシンが視床下部から分泌される。 - 3.鋤鼻器
フェロモンを受容する、嗅覚系の器官。 - 4.分界条床核BST
広義の扁桃体に属する終脳の領域で、不安やストレス応答に関与していると考えられている。 - 5.内側視索前野中央部cMPOA
終脳底部に属する内側視索前野の中央部。恒岡・黒田らにより、マウス母性行動にもっとも重要な働きをする場所として同定された(2012)。 - 6.光遺伝学的手法
光感受性タンパク質(後述のチャネルロドプシン等)を、遺伝学を用いて特定の神経細胞群に発現させ、その神経細胞群に局所的に光を当てて活性化させたり、抑制したりする技術。 - 7.フェロモン
動物が体内で生成して体外に分泌すると、同種の他の個体に対して、一定の行動を促す生理活性物質のこと。 - 8.狭鼻猿類
ユーラシア・アフリカに生息する霊長類で、ヒヒやラングール、類人猿などを含む。 - 9.c-Fosタンパク質
最初期遺伝子群のひとつである cFOS遺伝子によってコードされるたんぱく質分子で、転写因子として働く。c-Fosの発現量は神経細胞が活動し転写活性が上昇した時にその程度に応じて上昇するため、神経細胞の活動・活性化の指標(マーカー)としてよく用いられる。 - 10.真陽性率、偽陽性率
本実験の場合、真陽性率とは、実際の行動が養育(または攻撃・単独)のマウス数のうち、正しくモデルにより行動が推定されたマウス数の割合を指す。偽陰性率とは、実際の行動が養育(または攻撃・単独)ではなかったマウス数のうち、誤ってモデルによって養育群(または攻撃・単独群)と推定されたマウス数の割合である。 - 11.GABA作動性の抑制性神経
他の神経細胞の活動を抑える働きをする神経細胞で、γ—アミノ酪酸(GABA)を伝達物質として機能する神経細胞。 - 12.チャネルロドプシン
緑藻類クラミドモナスから同定された、光活性化非選択的陽イオンチャネルで、470nmの青色光照射によって神経細胞を活性化する性質をもつ。
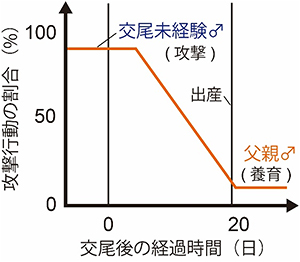
図1 父親マウスになる際の行動変化
交尾未経験オスマウスは、高確率で子を攻撃する。しかし、交尾・共生を経てメスマウスが出産をすると、オスマウスの攻撃行動は低下し、父性養育行動をとるようになる。
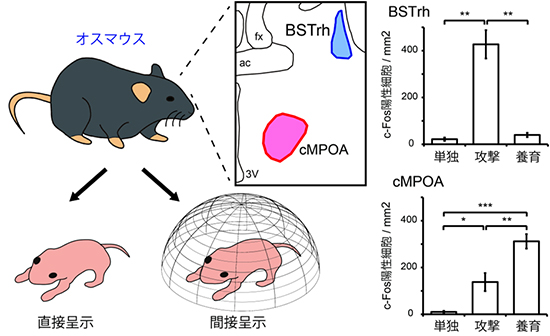
図2 養育行動と攻撃行動に関連する脳領域の同定
オスマウスの脳の活性化部位を、c-Fosタンパク質を指標としてみた結果、攻撃行動でBSTrhが、養育行動でcMPOAがそれぞれ活性化することが示された。cMPOAは攻撃行動時にもある程度活性化するが、養育行動時ほど顕著ではない。これらの結果は、子に金網をかけて、実際に攻撃行動や養育行動ができない状況にしても基本的に同様だった(図中の結果は直接呈示群のみ)。対照群は、2時間単独でいたオスマウス(単独)。
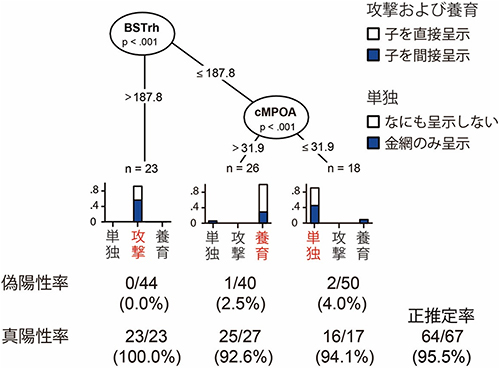
図3 樹木モデルによる行動推定とその精度
BSTrhとcMPOAのc-Fos陽性細胞数を定量化し解析をおこなうと、95%以上という高い精度で攻撃、養育、単独飼育の3種類の行動を推定できた。また、行動の推定は、子を直接呈示(白)するか、金網を使って間接的に呈示(青)するかという方法の違いには影響されなかった。
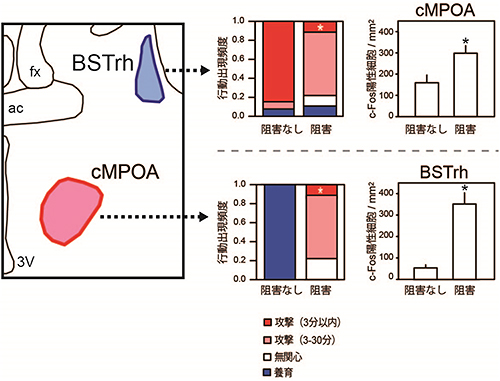
図4 BSTrhとcMPOAの阻害実験
交尾未経験オスのBSTrhを阻害すると、阻害なしの群と比べて、より多くのオスマウスにおいて子への攻撃を始めるまでの時間が長くなる(濃赤から薄赤)。さらに、BSTrhの阻害によってcMPOAのc-Fos陽性細胞数が増加したことから、cMPOAが活性化していることが示された。一方、父親マウスのcMPOAを阻害すると、養育(青)するオスマウスは皆無になり、攻撃(濃赤と薄赤)するオスマウスが有意に増えた。
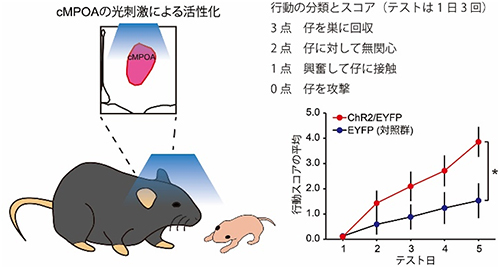
図5 cMPOAの活性化による交尾未経験オスの攻撃行動の減少
cMPOA特異的にチャネルロドプシンを発現させ、青色光を当ててcMPOA神経細胞を活性化させると、攻撃行動が減少し、実験開始5日後には養育行動スコアが有意に増加した。
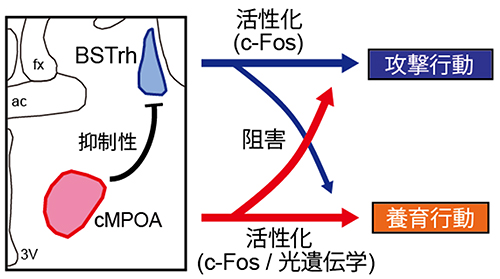
図6 父性養育行動に関わる神経回路
BSTrhは攻撃行動を促進しcMPOAは養育行動を促進するという、互いに相反する機能を持つ。cMPOAは抑制性の投射をBSTrhにのばしBSTrhの働きを抑えている。一方BSTrhもcMPOAを阻害する機能を示すが限定的であり、間接的なメカニズムであることが考えられる。