要旨
理化学研究所(理研)脳科学総合研究センター精神生物学研究チームの内匠透シニアチームリーダー、仲西萌絵リサーチアソシエイトらの国際共同研究グループ※は、自閉症(自閉スペクトラム症)の発症に関与する可能性がある遺伝子として、新たに「NLGN1」を同定しました。
自閉症は、社会的コミュニケーションの障害と繰り返し行動、こだわりの強さを特徴とする発達障害の一つです。自閉症は遺伝的要因が強く関与する疾患であり、特定の遺伝子の変異が発症の原因となることが知られています。なかでも近年、シナプス[1]に関連する遺伝子の変異が多く同定されていることから、シナプス異常と自閉症の関連が強く示唆されています。
今回、共同研究グループは、自閉症患者から新たにNLGN1と呼ばれるシナプス関連遺伝子の変異を同定し、患者で発見された変異がNLGN1タンパク質の減少などの異常を引き起こしていること、またシナプス形成不全などの機能喪失を引き起こしていることを明らかにしました。さらに、患者で発見された変異を導入したモデルマウスを作製したところ、変異を持ったマウスは社会的コミュニケーションや空間記憶能力などに異常を示すことが分かりました。
これまでに、NLGN-NRXN-Shank経路[2]では、NLGNファミリーのNLGN3やNLGN4が自閉症関連遺伝子として同定されていました。本成果は、自閉症におけるこの経路の重要性を裏付けるものです。今後、NLGN-NRXN-Shank経路を標的とした治療薬の開発や自閉症メカニズムの解明への貢献が期待できます。
本研究は、米国のオープンアクセス科学雑誌『PLOS Genetics』(8月25日付け:日本時間8月26日)に掲載されます。
※国際共同研究グループ
理化学研究所 脳科学総合研究センター
精神生物学研究チーム
シニアチームリーダー 内匠 透(たくみ とおる)
リサーチアソシエイト 仲西 萌絵(なかにし もえ)
研究員 野村 淳(のむら じゅん)
研究員 玉田 紘太(たまだ こうた)
動物資源開発支援ユニット
支援ユニットリーダー 高橋 英機(たかはし えいき)
テクニカルスタッフ 荒井 高(あらい たかし)
ペンシルベニア大学 医学大学院
教授 マヤ・ブカン(Maja Bućan)
大学院生(研究当時) シャオ・ジ(Xiao Ji)
背景
自閉症(自閉スペクトラム症)は、社会的コミュニケーションの障害と繰り返し行動、特定のものへのこだわりの強さを特徴とする発達障害の一つです。
自閉症の発症には、遺伝的要因が大きく関与することが知られており、さまざまな遺伝子変異が発症に関与すると考えられていますが、原因となる遺伝子の全容と詳しい分子メカニズムは明らかでありません。近年、シナプスに関連する遺伝子の変異が多く同定されていることから、シナプス異常と自閉症の関連が強く示唆されています。
シナプス膜に存在する分子(タンパク質)のNeuroligin(NLGN)と、その結合分子であるNeurexin(NRXN)、Shankから構成されるNLGN-NRXN-Shank経路では、これまでに複数の自閉症関連遺伝子が同定されており、この分子経路の異常は自閉症の発症と強く関連していると考えられます。これまでにNLGNファミリーのNLGN3やNLGN4が自閉症関連遺伝子であることが明らかにされてきましたが、他の分子の関与は明らかではありませんでした。
研究手法と成果
国際共同研究グループはまず、自閉症の兄弟2名の全エクソン(ゲノムのうちタンパク質をコードしている部分)の塩基配列を解読し、兄弟が共通して持っているミスセンス変異[3]を全て抽出しました。その中で、健常者のデータベース(約60,000人)に発見されず、さらにコンピュータプログラムによる機能変化予測解析で危険度が高いと予測されたのが、NLGN1上にある一つのミスセンス変異でした(図1A)。国際共同研究グループは、他の自閉症患者でもNLGN1の変異を持つケースがあるのではないかと考え、過去に発表された患者のデータ(約2500家族)に加えて未発表のデータ(患者362人)も調べ、さらに5人の患者が四つのNLGN1上にミスセンス変異を持つことが分かりました。
次に、実際に自閉症患者で発見されたNLGN1上のミスセンス変異が、NLGN1タンパク質の機能変化を引き起こしているかを調べるために、培養細胞を用いた発現実験を行いました。その結果、同定した五つの変異型タンパク質のうち四つでは、正常タンパク質に比べて発現量が減少していました(図1B)。さらに、発現量が減少した四つの変異型タンパク質のうち三つでは、細胞内での局在が小胞体[4]に限局するなど異常なパターンを示しました(図1C)。また、五つの変異型タンパク質のうち一つは、NLGN1タンパク質が分解される過程で異常を示していることが明らかとなり、NLGN1タンパク質の減少は少なくとも二つ以上の異常な経路を通じて引き起こされていることが分かりました。さらに、初代培養神経細胞を用いた発現実験を行ったところ、正常なNLGN1の持つシナプス後部構造のスパイン[5]形成機能が、変異型のタンパク質では消失していました。これらの結果から、自閉症患者で同定された変異は、NLGN1タンパク質の発現や機能に異常を引き起こすことが明らかになりました。
続いて、同定したNLGN1遺伝子変異やそれによって引き起こされるNLGN1タンパク質の異常が、実際に自閉症に特徴的な行動変化をもたらすのかを解析しました。CRISPR/Cas9[6]と呼ばれるゲノム編集技術を用いて、患者で同定されたものと同じミスセンス変異を持った患者モデルマウスであるNlgn1遺伝子変異マウス(変異型マウス)を作製し、その行動を観察しました。マウスの社会性を評価するために、空のケージとケージに入ったマウスをそれぞれ提示し、変異型マウスがアプローチして匂いを嗅ぐなどする時間の測定を行いました。その結果、変異型マウスでは、空のケージにアプローチする時間は野生型と変わりませんでしたが、ケージに入ったマウスにアプローチする時間は、野生型マウスと比較して短いことが分かりました(図1D上)。自閉症患者は、社会的コミュニケーションの障害を示しますが、この結果は変異型マウスにおける社会的興味の低さや社会的コミュニケーションの異常を示していると考えられます。また、変異型マウスは、空間記憶の能力にも異常があることが分かりました(図1D下)。
以上の結果は、NLGN1上のミスセンス変異がNLGN1タンパク質の減少など、タンパク質レベルでの異常を引き起こすこと、さらにその異常が自閉症様の行動異常につながることを示していることから、NLGN1は新たな自閉症候補遺伝子であるといえます。
今後の期待
これまでに、NLGNとその結合分子のからなるNLGN-NRXN-Shank経路の異常は、自閉症の発症において重要であると考えられてきました。本成果はその仮説を裏付けるものです。今後、ヒトの患者と同じ変異を持ったマウスの解析を行うことで、NLGN-NRXN-Shank経路を標的とした治療薬の開発や自閉症のメカニズムの解明につながると期待できます。
原論文情報
- Moe Nakanishi, Jun Nomura, Xiao Ji, Kota Tamada, Takashi Arai, Eiki Takahashi, Maja Bućan, Toru Takumi, "Functional significance of rare neuroligin 1 variants found in autism", PLOS Genetics, doi: 10.1371/journal.pgen.1006940
発表者
理化学研究所
脳科学総合研究センター 精神生物学研究チーム
シニアチームリーダー 内匠 透(たくみ とおる)
リサーチアソシエイト 仲西 萌絵(なかにし もえ)
 内匠 透
内匠 透
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
お問い合わせフォーム
産業利用に関するお問い合わせ
理化学研究所 産業連携本部 連携推進部
お問い合わせフォーム
補足説明
- 1.シナプス
神経細胞同士の情報伝達に関わる構造。情報を伝える細胞と伝えられる細胞の間には約20ナノメートル(1nmは10億万分の1m)のすき間がある。情報を伝える細胞はこのすき間に神経伝達物質を放出し、伝えられる細胞側の神経伝達物質受容体がそれを受け取ることにより、神経情報が伝わる。 - 2.NLGN-NRXN-Shank経路
NLGN、NRXNは、それぞれリガンド-受容体の関係でシナプス膜に発現するシナプス形成に関わるタンパク質、Shankは後シナプスに存在するタンパク質である。これらのタンパク質の変異が自閉症の原因(感受性)遺伝子としても知られており、本経路は自閉症を含む精神疾患に重要なシグナル経路と考えられる。 - 3.ミスセンス変異
一塩基置換変異のうち、タンパク質を構成するアミノ酸の置換を伴う変異のこと。 - 4.小胞体
細胞小器官の一つ。分泌経路にのるタンパク質を合成する場で、N型糖鎖の付加やタンパク質の正しい折り畳みが行われる。 - 5.スパイン
樹状突起フィロポディアが成熟したシナプス後部の構造で、その多くはキノコのような形態をしている。哺乳類では、興奮性情報伝達の90%以上を担っている。てんかんや脆弱性X症候群などの脳疾患において、スパイン形態の異常が報告されている。 - 6.CRISPR/Cas9
CRISPR/Cas系を用いたゲノム編集技術の一つ。CRISPR/Cas系は古細菌が持つ獲得免疫機構で、外来性のDNAを自らのゲノムに組み込み、2度目の感染のときにこの外来性のDNAの断片から二つのガイドRNAを発現させ、侵入してきた標的DNAを、Cas9ヌクレアーゼを用いて切断する系である。一つのガイドRNAとCas9の二つの因子だけでゲノムDNAの切断ができることが分かっており、手法の簡便さからゲノム編集の主たる手法となりつつある。
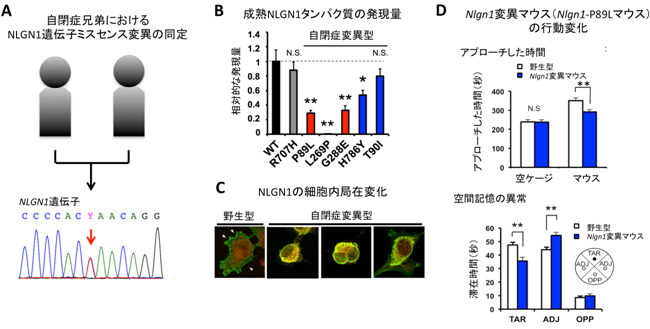
図1 自閉症患者におけるNLGN1遺伝子変異の同定と表現型解析
A)自閉症の兄弟2名で発見されたNLGN1遺伝子ミスセンス変異。7番目の塩基(Y)が健常者ではシトシン(C)のところが、自閉症患者ではチミン(T)に置換されていた(赤矢印の部分)。
B)発見された自閉症変異型NLGN1を過剰発現させ、発現量を解析したところ、自閉症患者で発見された五つの変異型(P89L、L269P、G288E、H786Y、T90I)のうち、T90Iを除く四つで発現量が減少していた。R707Hは一部の健常者でみられる変異型である。
C)細胞内局在解析では、野生型(WT)が細胞表面に強く発現するのに対し、三つの自閉症変異型(左からP89L、L269P、G288E)では小胞体に局在が変化していた。緑色がNLGN1タンパク質の発現、赤色が小胞体のマーカーを示している。
D)Nlgn1遺伝子変異マウスでは、空のケージにアプローチする時間は野生型と変わらなかったが、ケージに入った他のマウスにアプローチする時間が野生型と比較して減少していた(上)。モリス水迷路と呼ばれる試験において、Nlgn1遺伝子変異マウスは空間記憶能力に異常を示した(下)。