理化学研究所(理研)生命機能科学研究センター個体パターニング研究チームの濱田博司チームリーダー、梶川 絵理子テクニカルスタッフⅠ、分子配列比較解析チームの工樂樹洋チームリーダーらの共同研究グループは、動物の体の内部のつくりが左右非対称となる仕組みを爬虫類では初めて分子レベルで解明しました。
多くの動物の体は、表面的には左右対称ですが、体の内部構造のほとんどは非対称です。分子レベルの研究から、胚発生の初期に細胞の動く繊毛[1]がつくり出す水流が、Nodalタンパク質というシグナル分子[2]の左右軸に沿った濃度勾配を生み、これが心臓をはじめとする臓器の形や配置における左右非対称性をつくり出すことが知られています。この仕組みは、哺乳類、両生類、硬骨魚類で共通していますが、鳥類(ニワトリ)の胚には動く繊毛がないことから、脊椎動物の中で鳥類は例外扱いされてきました。
今回、共同研究グループは、理研でモデル生物としてのリソース化を進めてきた爬虫類ソメワケササクレヤモリとスッポンを用いて調べた結果、鳥類と同様に、爬虫類も繊毛に依存せずに左右非対称となる仕組みを持つことが分かりました。また、爬虫類と鳥類とで共通した仕組みは、脊椎動物が本来二つ持っているNodal遺伝子の使い分けが影響し、羊膜類[3]の進化の中で哺乳類を導いた系統から分岐した後、爬虫類と鳥類に至る系統で獲得されたことを示しました。
本研究は、英国のオンライン科学雑誌『Nature Ecology & Evolution』(1月6日付)に掲載されました。

図 (a)ソメワケササクレヤモリの成体と(b)初期発生において左右対称性が破れる仕組み
背景
多くの動物の体は表面的には左右対称ですが、例えばヒトの心臓のように、体の内部にある臓器のほとんどは、その形や配置が左右非対称です。この非対称性は、体ができる過程の早い段階で生まれ、各臓器の機能や臓器間の連携を保つために必須で、異常を来すと、多くの場合心臓の奇形が生じ、出生後に生命の危機に陥ります。
さまざまなモデル動物を用いた過去の研究から、左右非対称な臓器の形や配置ができる過程の一部は種々の生物で共通しているものの、残りの過程には多様性があることが分かっています。共通した役割を果たす重要なシグナル分子としてNodalタンパク質があり、このタンパク質は、胚発生の初期において左側に分布することが、マウス、アフリカツメガエル、ニワトリ、ゼブラフィッシュで共通して観察されています。
一方、異なる種間での多様性が最も目立つのは、胚発生の初期段階において左右の対称性が破れる仕組みです。脊椎動物の硬骨魚類、両生類、哺乳類では、胚の特定の細胞が持つ繊毛が回転運動し、周りの液体を左方向へ動かすことで、対称性を破ると考えられていますが、ニワトリ胚の細胞にはそのような動く繊毛がありません。しかし、なぜ鳥類だけが例外なのか、繊毛を使わずにどのようにして左右非対称となるのかは明らかになっていませんでした。
共同研究グループは今回、理研で全ゲノム解読[4]を行うなどモデル生物としてのリソース化が進められてきた爬虫類のソメワケササクレヤモリ注1,2)とスッポン注3)を対象として、爬虫類は対称性を破るのに繊毛を使うのか、使わないのであればどのような仕組みで非対称となるのか、という謎の解明に挑みました。
- 注1)2018年4月16日プレスリリース「爬虫類ソメワケササクレヤモリの全ゲノム解読」
- 注2)2015年11月20日プレスリリース「新たな実験動物としてのソメワケササクレヤモリ」
- 注3)2013年4月29日プレスリリース「ゲノム解読から明らかになったカメの進化」
研究手法と成果
左右非対称性の決定に重要な役割を果たすNodalタンパク質を作り出す遺伝子の数は、サメやシーラカンスでは二つ、ゼブラフィッシュでは三つ、ニワトリ、マウスでは一つと異なり、それらの遺伝子の系統関係は長らく整理されてきませんでした。
そこで共同研究グループは、多様な脊椎動物のゲノム[4]のDNA塩基配列全体について、綿密な分子系統解析を行いました。その結果、脊椎動物の共通祖先はおそらく二つのNodal遺伝子を持ち、それらの両方あるいは片方が受け継がれてきたことが示されました(図1)。共同研究グループはそれぞれの遺伝子を、「Nodal1」および「Nodal2」と命名しました。奇妙なことに、進化の過程で哺乳類は、Nodal2遺伝子をゲノムから失った結果Nodal1遺伝子だけを保持し、反対に多くの鳥類・爬虫類(ソメワケササクレヤモリとスッポン)は、Nodal1遺伝子を失った結果Nodal2遺伝子だけを保持していることが分かりました(図1)。
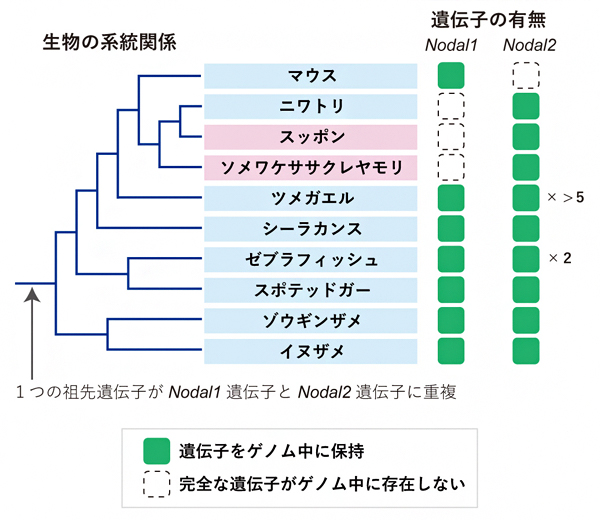
図1 脊椎動物のNodal遺伝子の重複、保持、消失の歴史
脊椎動物の共通祖先は二つのNodal遺伝子(Nodal1とNodal2)を持ち、進化の過程で哺乳類(マウス)はNodal2遺伝子を失った結果Nodal1遺伝子だけを保持し、鳥類(ニワトリ)、爬虫類(スッポン、ソメワケササクレヤモリ)はNodal1遺伝子を失った結果Nodal2遺伝子だけを保持している。また、ツメガエルは独自にNodal2に対応する遺伝子を六つ以上に増やし、ゼブラフィッシュはNodal2に対応する遺伝子をニつ保持している。
左右の対称性が破れる際、Nodal遺伝子はまず胚の左右オーガナイザー[5]で発現し、その後側板中胚葉[6]で発現します。側板中胚葉におけるNodal遺伝子の発現は、マウスNodal1遺伝子とニワトリNodal2遺伝子は共通して左側でのみ見られます。今回、ソメワケササクレヤモリとスッポンはニワトリと同様にNodal2遺伝子だけを持つことが分かったため、それらの左右オーガナイザーと側板中胚葉における遺伝子発現パターンをin situハイブリダイゼーション法[7]を用いて調べました。その結果、側板中胚葉では、どちらの種でも左側でのみNodal2遺伝子の発現が確認されました(図2)。
左右オーガナイザーにおけるNodal遺伝子の発現は種によって異なり、細胞に動く繊毛を持つマウス、ツメガエル、ゼブラフィッシュでは、発現の強さは左右で大きな差はありません。しかし、Nodalタンパク質の活性を抑制する物質が、繊毛の回転で生じる水流によって左側特異的に分解されるため、左側でNodalタンパク質の活性が強くなります注4)。
一方、動く繊毛を持たないニワトリでは、Nodal2遺伝子の発現強度がはじめから左右オーガナイザーの左側で強く、そのために左側のNodal2タンパク質の活性が強くなります。ソメワケササクレヤモリとスッポンの左右オーガナイザーでは、ニワトリと同様にNodal2遺伝子の発現ははじめから左側で強いことが分かりました(図2)。この結果から、爬虫類でも鳥類と同様に、左右対称性が破れる際に繊毛は関与しないと予測されました。
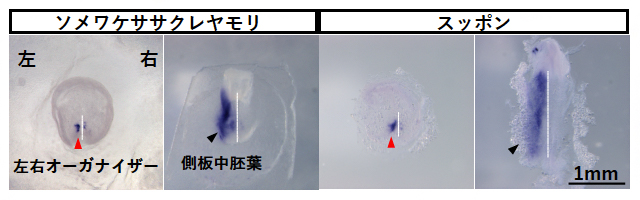
図2 ソメワケササクレヤモリおよびスッポンの胚におけるNodal2遺伝子の発現パターン
ソメワケササクレヤモリとスッポンそれぞれにおいて、左側が左右オーガナイザー、右側が側板中胚葉の写真。in situハイブリダイゼーションにより、Nodal2遺伝子の発現部位が紫色に染まっている。ヤモリとスッポンのどちらも、左右オーガナイザー(赤矢頭)および側板中胚葉(黒矢頭)において、左側のみNodal2遺伝子が発現していることが分かる。白線は左右を分ける軸を示す。
そこで、動く繊毛を持つ細胞で特異的に現れる遺伝子が、爬虫類の左右オーガナイザーにあたる原口の周辺に発現しているかを免疫染色法[8]およびin situハイブリダイセーション法で調べたところ、それらの遺伝子の発現は原口周辺では見られませんでした。また、電子顕微鏡およびライブイメージングによって、実際に動く繊毛があるかどうかも探索しましたが、動く繊毛は観察されませんでした。
次に、左右オーガナイザーにおけるNodal1遺伝子とNodal2遺伝子の発現パターンの違いがどのような仕組みで生じるかを明らかにするために、これら遺伝子の発現を制御するエンハンサー[9]の塩基配列の違いを調べました。その結果、側板中胚葉での遺伝子発現を制御するエンハンサー塩基配列は、調べた全ての種においてNodal1遺伝子とNodal2遺伝子の周辺に存在していました。一方、左右オーガナイザーでの発現を制御するエンハンサー塩基配列は、哺乳類ではNodal1遺伝子の周辺に存在したのに対し、爬虫類と鳥類のNodal2遺伝子の周辺には存在しませんでした。
そこで、これらエンハンサー塩基配列の違いが、左右オーガナイザーにおけるNodal1遺伝子とNodal2遺伝子の発現の違いをもたらすのかどうかを明らかにするために、マウスNodal1エンハンサーまたはニワトリNodal2エンハンサーでレポーター遺伝子[10]LacZを発現させるトランスジェニックマウス[11]をそれぞれ作製しました。その結果、それぞれのトランスジェニックマウス胚において、マウスNodal1エンハンサーで発現制御されたLacZ遺伝子は、左側の側板中胚葉と左右オーガナイザーの両側で発現したのに対し、ニワトリNodal2エンハンサーで発現制御されたLacZ遺伝子は、左側の側板中胚葉でのみ発現し、左右オーガナイザーでは発現しませんでした(図3)。
従って、側板中胚葉でNodal1遺伝子とNodal2遺伝子を発現させる仕組みは、哺乳類と鳥類・爬虫類で共通である一方、左右オーガナイザーでの発現は異なる仕組みによって制御されていることが示されました。
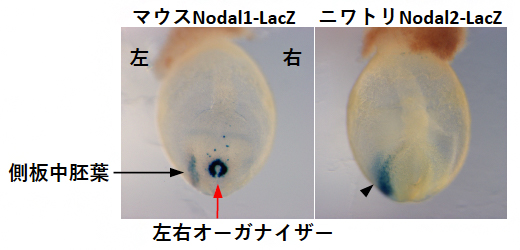
図3 トランスジェニックマウス胚におけるレポーター遺伝子LacZの発現パターン
- 左:マウスNodal1エンハンサーによって発現制御されたLacZの発現パターン。左側の側板中胚葉と左右オーガナイザーの両側で発現している。
- 右:ニワトリNodal2エンハンサーにより発現制御されたLacZの発現パターン。左右オーガナイザーでは発現せず、左側の側板中胚葉でのみ発現している。
- 注4)2017年4月4日CDBニュース「左右非対称性を生み出すWnt5のグラデーション」
今後の期待
共同研究グループでは、今後、鳥類や爬虫類が、なぜ動く繊毛なしで左右非対称性を成すことができるのか、鳥類や爬虫類の個体発生の過程における左右非対称性の起源は何なのかといった疑問に答えていきたいと考えています。
そのためにはまず、ニワトリNodal2遺伝子がニワトリ胚の左右オーガナイザーにおいて、非対称な発現をもたらすエンハンサーを探索・同定する必要があります。そのようなエンハンサーが分かれば、それを糸口にして、繊毛に頼らずに非対称性を成すメカニズムが解明でき、ひいては動物界における左右対称性を破る仕組みの多様性を理解できると考えています。
補足説明
- 1.繊毛
細胞から突出する微小管を含む構造(オルガネラ)。繊毛には動くものと動かないものがあり、前者には、気道上皮・脳室上衣の繊毛や精子の鞭毛がある。後者は体の中のほとんどの細胞に見られ、細胞外からの種々のシグナルを感知する役割を持つ。 - 2.シグナル分子
細胞内や細胞間で情報を伝える働きを持つ分子。その一つであるNodalタンパク質は、細胞間シグナル伝達に関わる分子の一つであるTGF-βファミリーに属し、細胞から分泌され、特定の受容体を介して他の細胞に情報を伝える働きをする。 - 3.羊膜類
胚発生の過程において、胚体を包む膜である羊膜を持つ動物。具体的には、現存の生物では爬虫類、鳥類および哺乳類を指す。 - 4.全ゲノム解読、ゲノム
「ゲノム」とは、種を特徴づける遺伝情報全体のことで、具体的には、細胞の核の中にある特定の数の染色体に納められたDNA分子一式を指す。ヤモリやスッポンなどの脊椎動物の場合、ゲノムにはタンパク質をつくる遺伝子が2万個以上存在する。生物種の間で遺伝子の数や多様性を正確に議論するためには、まずゲノム全体のDNA配列を明らかにする「全ゲノム解読」を行う必要がある。 - 5.左右オーガナイザー
左右対称性が破られる胚の部位。マウス胚では「ノード」、ニワトリ胚では「ヘンゼン結節」、爬虫類胚では「原口」と呼ばれる場所。 - 6.側板中胚葉
胚の周辺部に左右両側性に見られる中胚葉。やがて、さまざまな腹腔内臓器の間質を形成する。 - 7.in situハイブリダイゼーション法
組織や細胞中で(in situ:本来の場所で)、特定の配列を持つ核酸(ここでは転写産物であるRNA)の分布を調べる方法。目的の配列に相補的な配列を持つ核酸分子を生体内には存在しない分子で標識し、核酸が特定の条件下では相補的な塩基同士でのみ結合する(ハイブリダイゼーションまたは雑種形成法)性質を利用して、目的の配列の核酸を捕捉する。その後、標識に用いた分子を蛍光色素や発色剤を使用して検出することで、その分布を調べる実験法。生物種を問わず、大きく実験手順を変更することなく適用できる手法である。 - 8.免疫染色法
組織切片中の標的分子に対する抗体を使い、その組織内でのタンパク質の局在を可視化する手法。免疫組織化学染色法とも呼ばれる。 - 9.エンハンサー
遺伝子の転写効率を著しく高めるDNA配列。多くの場合、一つの遺伝子は、種々の異なる細胞で働く複数のエンハンサーを持つ。 - 10.レポーター遺伝子
ある遺伝子の発現や活性を簡単にモニターするための外来性遺伝子。例えば、大腸菌由来のLacZ遺伝子やクラゲ由来の緑色蛍光タンパク質のGFP遺伝子などがある。 - 11.トランスジェニックマウス
広義では何らかの遺伝子改変を持つマウスであるが、通常は狭義で用いられ、人工的な操作により外来性の遺伝子がゲノム中に挿入されたマウスを意味する。
共同研究グループ
理化学研究所 生命機能科学研究センター
個体パターニング研究チーム
チームリーダー 濱田 博司(はまだ ひろし)
テクニカルスタッフⅠ 梶川 絵理子(かじかわ えりこ)
研究員 井手 隆広(いで たかひろ)
研究員 水野 克俊(みずの かつとし)
研究員 峰岸 かつら(みねぎし かつら)
テクニカルスタッフⅠ 井川 弥生(いかわ やよい)
テクニカルスタッフⅠ 西村 博美(にしむら ひろみ)
分子配列比較解析チーム
チームリーダー 工樂 樹洋(くらく しげひろ)
基礎科学特別研究員(研究当時) 原 雄一郎(はら ゆういちろう)
生体モデル開発チーム
チームリーダー 清成 寛(きよなり ひろし)
大阪大学SEEDSプログラム(研究当時)
SEEDS2017実感コース受講生 保呂 有珠暉(ほろ うずき)
大阪大学大学院 生命機能研究科
助教 内川 昌則(うちかわ まさのり)
研究支援
本研究の一部は、科学技術振興機構(JST)戦略的創造研究推進事業CREST「生命動態の理解と制御のための基盤技術の創出(領域代表者:山本雅)」、日本学術振興会(JSPS)科学研究費補助金(基盤研究A)「形態の非対称性の起源(研究代表者:濱田博司)」による支援を受けて行われました。
原論文情報
- Eriko Kajikawa, Uzuki Horo, Takahiro Ide, Katsutoshi Mizuno, Katsura Minegishi, Yuichiro Hara, Yayoi Ikawa, Hiromi Nishimura, Masanori Uchikawa, Hiroshi Kiyonari, Shigehiro Kuraku, Hiroshi Hamada, "Nodal paralogues underlie distinct mechanisms for visceral left-right asymmetry in reptiles and mammals", Nature Ecology & Evolution, 10.1038/s41559-019-1072-2
発表者
理化学研究所
生命機能科学研究センター 個体パターニング研究チーム
チームリーダー 濱田 博司(はまだ ひろし)
テクニカルスタッフⅠ 梶川 絵理子(かじかわ えりこ)
生命機能科学研究センター 分子配列比較解析チーム
チームリーダー 工樂 樹洋(くらく しげひろ)
 濱田 博司
濱田 博司
 工樂 樹洋
工樂 樹洋
報道担当
理化学研究所 広報室 報道担当
お問い合わせフォーム