2020年6月26日
理化学研究所
シンガポール国立大学メカノバイオロジー研究所
筑波大学
細胞の追いかけっこが波を作る
-細胞の追随運動が細胞密度を伝搬させるメカニズムを解明-
理化学研究所(理研)生命機能科学研究センターフィジカルバイオロジー研究チームの柴田達夫チームリーダー、早川雅之研究員、シンガポール国立大学メカノバイオロジー研究所の平岩徹也特別研究員(MBI Fellow)、筑波大学生命環境系の桑山秀一准教授らの国際共同研究グループは、「密度波[1]」として現れる細胞性粘菌[2]細胞の集団運動が、「接触追随[3](Contact following locomotion)」という一過性の単純な細胞間相互作用により引き起こされることを明らかにしました。
接触追随は哺乳類細胞でも観察される現象であり、本研究成果は、多細胞生物の形態形成における細胞の複雑な集団運動の解明に貢献すると期待できます。
真核生物である細胞性粘菌は、単細胞期と多細胞期の生活環を持っています。化学物質の濃度勾配に沿って移動する走化性[4]に従って、一つ一つの細胞が集合し、多細胞体制を形成する過程は、細胞運動や形態形成のモデルとして広く研究されています。
今回、国際共同研究グループは、走化性を失った細胞性粘菌変異株が示す進行波[5]状の集団運動に着目し、遺伝学的手法、定量的解析や数理モデルを組み合わせることで、この集団運動の形成メカニズムの解明に取り組みました。その結果、進行波状の集団運動の正体は、細胞が集まっている部分とまばらな部分が動的に入れ替わることで生じる細胞密度の伝搬(密度波)であり、細胞が他の細胞に追随する接触追随という細胞間相互作用が密度波の維持に重要な役割を果たしていることを明らかにしました。
本研究は、オンライン科学雑誌『eLife』(4月30日付)に掲載されました。
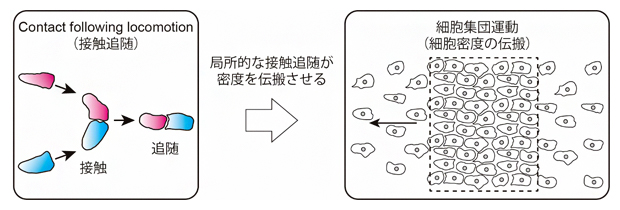
細胞の追随運動が集団運動を生み出す
背景
真核細胞の集団運動は、形態形成や創傷治癒など多細胞体制の構築・維持のプロセスで重要な役割を果たしており、生命科学の広い分野で研究されています。細胞の集団運動は一般的に、一部の細胞が分泌する化学物質の濃度勾配に沿って移動する「走化性」により他の細胞が誘導されたり、複数の細胞がある分子を介して強固に接着しながら一緒に運動したりすることで実現されます。例えば、細胞集団運動を示すモデル生物としてよく用いられる細胞性粘菌(Dictyostelium discoideum)は、通常、細胞が分泌するcAMP[4](環状アデノシン一リン酸)というシグナル分子への走化性によって集団運動が実現されます。つまり、細胞間における分子の放出と受容という高度な相互作用が集団運動を引き起こしています。
一方、近年の物理学における自己駆動粒子[6]の研究において、「隣の個体と運動方向をそろえる」という単純な相互作用によって、集団運動が可能になることが分かってきました。実際に、動物、細菌、生体高分子などさまざまなスケールの集団において、単純な相互作用による集団運動が観察されており、集団運動と個体・分子の挙動の関係性が明らかになっています。例えば、分子モーター[7]によって駆動される生体高分子が密集すると、個々の生体高分子同士の衝突という単純相互作用により、結果としてお互いが整列し、進行波状の集団運動が引き起こされます。
また、同様の進行波状の集団運動は、走化性を失った細胞性粘菌においても起こることが筑波大学の研究チームにより発見されています注1)が、集団運動と個々の細胞の挙動との関係性は不明のままでした。進行波状の集団運動の特徴付けや、原因となる細胞間相互作用を特定できれば、真核細胞が示す複雑な集団運動を理解する新たな鍵となる可能性があります。
- 注1)Kuwayama H, Ishida S. 2013. Biological soliton in multicellular movement. Scientific Reports 3:2272.
研究手法と成果
走化性を示さない細胞性粘菌の変異株(KI細胞[4])の集団では、野生株とは異なる進行波状の集団運動が観察されます。国際共同研究グループは、この集団運動と細胞間相互作用を定量的に解析し、形成メカニズムを実験・数値シミュレーション両側面から明らかにすることを試みました。
KI細胞を均一に寒天培地上に撒いて一定時間経過させると、細胞密度の低い領域と高い領域とに分かれます。高密度領域は帯状の構造を持っており、この帯に対して垂直方向に運動するため、波が進むように見えます(図1)。高密度領域の運動速度は、異なるサンプル間においてもほぼ一定で、毎秒約0.5マイクロメートル(μm、1μmは1,000分の1mm)でした。

図1 KI細胞の集団運動
寒天培地の平面上で、細胞の高密度領域が現れ、その領域が一方向(左方向)に運動する。
一方、高密度領域と低密度領域に存在する個々のKI細胞の運動速度は、それぞれ約0.4μm/sと約0.3μm/sであり、集団内のKI細胞一つ一つの速さが、KI細胞集団が形成する進行波そのものよりも遅いことが判明しました。また、1細胞を追跡した運動軌跡からは、細胞は低密度領域から高密度領域に移動して集団運動した(進行波として振る舞った)後、再び低密度領域に戻っていることが判明しました(図2左)。これらの観察から、KI細胞の進行波状の集団運動は、細胞が集まっている部分とまばらな部分ができ、それが動いていくことで生じる高密度領域の伝搬、すなわち「密度波」であることが分かりました(図2右)。
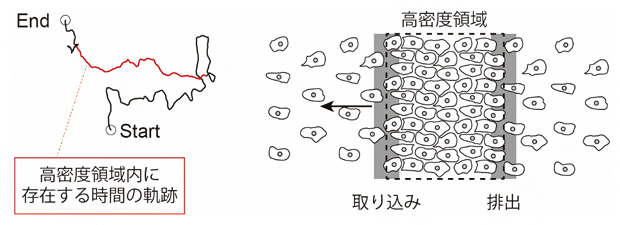
図2 KI細胞の運動解析
- (左)KI細胞の運動軌跡。黒線は細胞が低密度領域内に存在した時間の運動軌跡、赤線は細胞が高密度領域内に存在した時間の運動軌跡を示す。細胞は低密度領域から高密度領域に移動して集団運動した後、再び低密度領域に戻っていた。
- (右)密度波状集団運動の概念図。はじめ低密度領域にあった細胞は、いったん高密度領域内に取り込まれ、集団運動した後、再び高密度領域から出て低密度領域に戻る。
KI細胞の運動軌跡を詳しく見ると、細胞は高密度領域にいる間は波の進む方向に動き、低密度領域にいる間はさまざまな方向に動く傾向がありました。そこで、細胞集団の前後方向(細胞極性)がどれだけそろっているかを定量的に解析するため、秩序変数(Order parameter)[8]と呼ばれる評価値を、高密度領域の内と外に関してそれぞれ算出しました。その結果、秩序変数は高密度領域外で低い値を示す一方で、高密度領域内では高い値を示しました。これより、高密度領域内の細胞の極性は周りとそろっている、言い換えると集団は比較的同じ方向に動いていることが分かりました。つまり、高密度領域内には秩序状態が存在しているということです。
さらに、伝搬する高密度領域の細胞運動をより詳しく調べるために、オプティカルフロー[9]と呼ばれる手法を用いて解析を行った結果、高密度領域の内部において、運動方向の異なる二つの領域が存在することが分かりました(図3)。寒天培地上の平面において波の進行方向を180°とした場合、左斜め下(約220°方向)に運動する集団と左斜め上(約140°方向)に運動する集団とが、交互に存在するような内部構造が見られました。
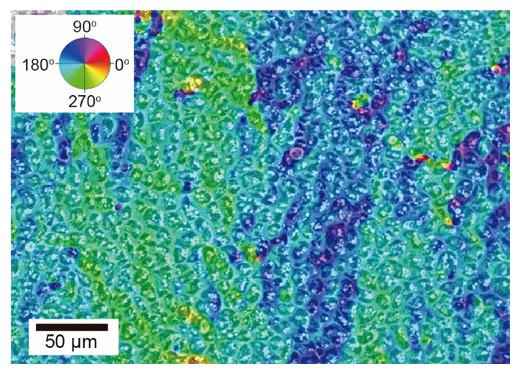
図3 高密度領域内の細胞構造
オプティカルフローと呼ばれる手法により、細胞が動いた角度に応じて色付けがされている。波が180°方向に進むとき、高密度領域内部には約220°方向に運動する集団(黄緑)と、約140°方向に運動する集団(青)とが交互に存在していた。
一般的に、高密度領域内にあるような秩序状態を生み出すには、要素間の相互作用が必要です。そこで、密度波状の集団運動の原因となっている細胞間相互作用を検証するために、細胞を1対1で衝突させ、どのような挙動を示すのか観察しました。その結果、衝突後に一方の細胞がもう一方の細胞に追随するような現象が見られ、国際共同研究グループはこの現象を「接触追随(Contact following locomotion;CFL)と名付けました(図4)。
CFLは野生株の細胞性粘菌においても観察され、また、これを引き起こす原因の一つとしてTgrB1[10]という細胞間接着を担うタンパク質が知られています。そこで、tgrb1遺伝子を発現できないKI細胞変異株を用いて、密度波状の集団運動が形成されるかどうか確かめたところ、形成されませんでした。したがって、密度波状の集団運動は、細胞間におけるCFLに起因するものであると結論付けられました。
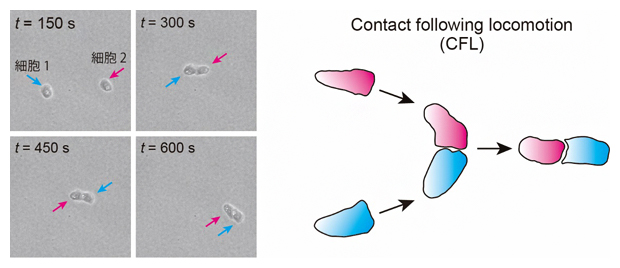
図4 接触追随(Contact following locomotion;CFL)
- (左)KI細胞の二体衝突。
- (右)接触追随の概念図。細胞を1対1で衝突させると、一方の細胞がもう一方の細胞に追随するような現象が見られた。
さらに、CFLの効果を取り入れた数理モデルを構築し、追随の相互作用により密度波状の集団運動が自発的に生じ得るのかどうかを調べました。構築したモデルを用いてシミュレーションを行った結果、ある条件ではCFLの強さに依存して密度波状の集団運動が形成されることが分かりました。また、高密度領域内に観察された運動方向の異なる領域も、細胞性粘菌での実験と同様に観察されました(図5)。
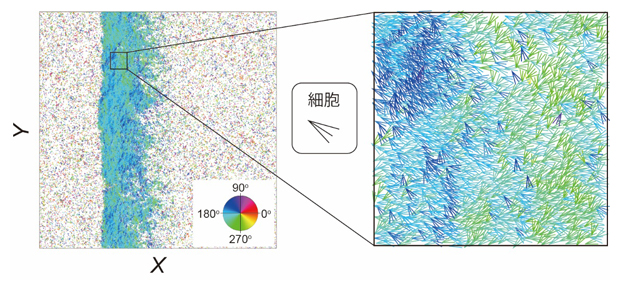
図5 CFLの効果を取り入れた数理モデルを用いたシミュレーション
- (左)シミュレーションによる密度波状の集団運動。
- (右)左の一部分を拡大したもの。細胞一つ一つが矢印で示され、その色は運動方向に対応している。図3と同様に、運動方向の異なる領域(色が異なる領域)が存在する。
今後の期待
本研究では、走化性を示さない細胞性粘菌変異株を用いて、CFLという細胞の追随相互作用が密度波状の集団運動を引き起こすことを実験と数理モデルで実証しました。自己駆動粒子による集団運動の形成という考え方が、細胞集団にも適用できることを示した成果です。
CFLは哺乳類の培養細胞でも観察されており注2)、真核細胞が持つ普遍的な性質である可能性があります。本研究で得られた知見から、CFLを示す哺乳類細胞の集団が細胞性粘菌と同様に密度波状の集団運動を引き起こすのか、またCFLは哺乳類の形態形成にどのような役割を果たしているのかという新たな問いが浮かび上がります。今後、さまざまな生物現象において、CFLの役割が明らかになっていくと期待できます。
- 注2)Li D, Wang YL. 2018. Coordination of cell migration mediated by site-dependent cell-cell contact. PNAS 115: 10678-10683.
補足説明
- 1.密度波
密度の違いが伝搬する波。疎密波とも呼ぶ。 - 2.細胞性粘菌
和名キイロタマホコリカビ。真核生物であり原生生物界に属する。森林土壌などに生息する。生活環において単細胞期と多細胞期がある。栄養が豊富な環境では単細胞のアメーバとして増殖するが、栄養飢餓に陥るとアメーバ細胞が集合して多細胞期が開始する。単細胞アメーバは哺乳類の白血球細胞と同じようにアメーバ運動するため、細胞運動や走化性のモデル生物として古くから利用されてきた。 - 3.接触追随
これまで、運動する細胞同士が衝突したときに進行方向を変えて離れる「接触阻害」現象はよく知られていた。これに対し本研究で見いだされた、衝突後に一方の細胞がもう一方の細胞に追随する現象を接触追随(Contact following locomotion;CFL)と名付けた。 - 4.走化性、cAMP、KI細胞
生物が化学物質の濃度勾配に沿って移動することを「走化性」と呼ぶ。細胞性粘菌は栄養飢餓になると自ら「cAMP(環状アデノシン一リン酸)」というシグナル分子を産生、分泌し、これに対して走化性を示すことで細胞が集合し多細胞体の形成が始まる。「KI細胞」は、cAMPに対する走化性を失った細胞性粘菌の変異株で、1993年に桑山秀一(当時京都大学)らによって単離された。 - 5.進行波
空間内をある方向に進む波。 - 6.自己駆動粒子
エネルギーを利用して自発的に推進する粒子。例えば、遊走する細胞も自己駆動粒子と見なすことができる。 - 7.分子モーター
細胞内でアデノシン三リン酸(ATP)の分解で生じる化学エネルギーなどを用いて、力学的な仕事をする分子の総称。筋肉の運動を担うミオシンや、繊毛・鞭毛の動きを担うダイニンなどがある。自己駆動粒子の実験では、ミオシンとアクチン、ダイニンと微小管の組み合わせなどを用いた。 - 8.秩序変数(Order parameter)
着目している系の持つ秩序を表す変数のこと。例えば本研究の場合、秩序状態では、秩序変数は高い値を示し、細胞が同じ方向に動いていることに対応する。また無秩序状態では、秩序変数は低い値を示し、集団内の細胞がそれぞれバラバラな方向に動いていることに対応する。 - 9.オプティカルフロー
時間的に連続している画像間において特徴的な点を探し出し、その変化から物体などの移動速度を推定する方法。 - 10.TgrB1
細胞性粘菌の持つ細胞接着分子の一つ。
国際共同研究グループ
理化学研究所 生命機能科学研究センター フィジカルバイオロジー研究チーム
チームリーダー 柴田 達夫(しばた たつお)
研究員 早川 雅之(はやかわ まさゆき)
(日本学術振興会(JSPS) 特別研究員(研究当時))
テクニカルスタッフ(研究当時) 和田 裕子(わだ ゆうこ)
シンガポール国立大学 メカノバイオロジー研究所
特別研究員(MBI Fellow) 平岩 徹也(ひらいわ てつや)
筑波大学 生命環境系
准教授 桑山 秀一(くわやま ひでかず)
研究支援
本研究は、日本学術振興会(JSPS)特別研究員奨励費「細胞性粘菌が示す集団運動の自発的な方向の非対称化、及びそれを用いた非対称構造形成(研究代表者:早川雅之)」による支援を受けて行われました。
原論文情報
- Masayuki Hayakawa, Tetsuya Hiraiwa, Yuko Wada, Hidekazu Kuwayama, and Tatsuo Shibata, "Polar pattern formation induced by contact following locomotion in a multicellular system", eLife, 10.7554/eLife.53609
発表者
理化学研究所
生命機能科学研究センター フィジカルバイオロジー研究チーム
チームリーダー 柴田 達夫(しばた たつお)
研究員 早川 雅之(はやかわ まさゆき)
シンガポール国立大学 メカノバイオロジー研究所
特別研究員(MBI Fellow) 平岩 徹也(ひらいわ てつや)
筑波大学 生命環境系
准教授 桑山 秀一(くわやま ひでかず)
 柴田 達夫
柴田 達夫
 早川 雅之
早川 雅之
 平岩 徹也
平岩 徹也
 桑山 秀一
桑山 秀一
報道担当
理化学研究所 広報室 報道担当
お問い合わせフォーム
筑波大学広報室
Email: kohositu [at] un.tsukuba.ac.jp
Tel: 029-853-2039 / Fax: 029-853-2014
※[at]は@に置き換えてください。