理化学研究所(理研)環境資源科学研究センター 植物免疫研究グループの白須 賢 グループディレクター(環境資源科学研究センター 副センター長)、ブルーノ・ポクマン・ゴウ 基礎科学特別研究員、門田 康弘 専任研究員らの国際共同研究チームは、植物の免疫受容体[1]の進化の軌跡を解明しました。
本研究成果は、植物免疫の起源と進化の理解につながるとともに、得られた知見を用いることで、植物のゲノム情報から免疫受容体として働く遺伝子と発生・成長に関わる遺伝子を迅速かつ正確に予測できるようになりました。
国際共同研究チームは、公開されている350種の植物ゲノム情報から、細胞膜に局在する受容体をコードする遺伝子を約21万個抽出して比較解析を行いました。そして、病原体の侵入を認識する免疫受容体群(ロイシンリッチリピート(LRR)[2]受容体型リン酸化酵素(LRR-RLKs)[3]、およびロイシンリッチリピート受容体様タンパク質(LRR-RLPs)[4])の進化の軌跡を調べました。その結果、LRR-RLPs型の免疫受容体群は発生・成長の制御を担う受容体群と共通の祖先から派生し、進化の過程でそれぞれの機能に必要なモジュール[5]を獲得することで異なる受容体へと進化したことが分かりました。
本研究は、科学雑誌『Nature Communications』オンライン版(2月1日付:日本時間2月1日)に掲載されました。
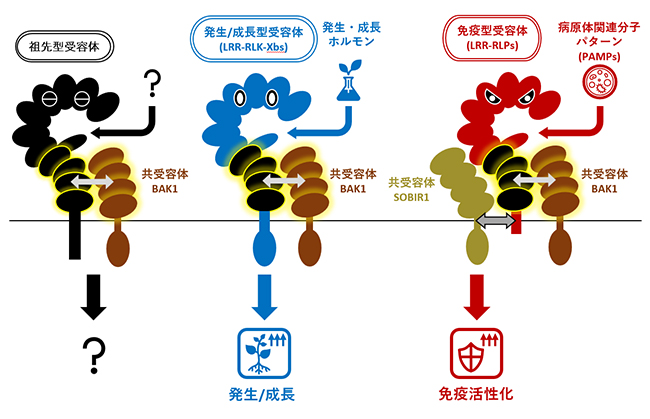
免疫に働く受容体と発生・成長を担う受容体は共通の祖先から派生し、共受容体BAK1と結合する領域を共有する
背景
植物が陸上に進出し、多様な微生物と遭遇する中で、どのようにして病原微生物を認識する能力を獲得したのか、植物免疫の起源に関しては未解明でした。
植物は細胞膜に微生物由来の物質を認識する受容体や、植物ホルモンを認識する受容体を持ち、この受容体は病原体の侵入を識別したり、植物細胞間での情報のやり取りをしたりします。このような受容体が細胞外からの情報を細胞内へと伝達していく過程において、タンパク質のリン酸化が重要な役割を果たします。一般に、細胞膜上の受容体はリガンド[6]を認識すると、受容体の中にあるアミノ酸がリン酸化され、活性化されます。そして、受容体の活性化は細胞内のタンパク質のリン酸化を誘導し、細胞の中へと情報が伝達されます。これら細胞膜局在型の受容体には、細胞質側にリン酸化酵素領域[7]を持つ受容体型リン酸化酵素(RLK)と、細胞質側に機能的な領域を持たない受容体様タンパク質(RLP)があります。この二つの受容体は細胞外領域の構造の違いによってさまざまなファミリーがあります。中でもLRR-RLPsとLRR-RLKsは植物において重要な役割を担っており、ロイシンリッチリピート(LRR)と呼ばれる繰り返し構造を介して、微生物由来の物質や植物ホルモンなど、さまざまなリガンドを認識し細胞内へと情報を伝えます。
免疫受容体として機能するLRR-RLPsは病原体に保存された病原体関連分子パターン(PAMPs)[8]を認識して免疫反応を誘導します。この免疫型LRR-RLPsは、共受容体[9]と呼ばれるタンパク質と協調して免疫反応を誘導します。免疫型LRR-RLPsは定常状態から共受容体SOBIR1[10]と結合しており、PAMPsを認識するともう一つの共受容体BAK1[11]と結合します。これにより、SOBIR1とBAK1のリン酸化酵素領域が近づくことで互いに活性化し、協調して下流因子をリン酸化することで情報を伝達して免疫反応を誘導します。一方、LRR-RLKsは細胞質側にあるリン酸化酵素領域を介して細胞内へと情報を伝達します。このリン酸化酵素領域のアミノ酸配列に従って、LRR-RLKsは機能別に20のグループ(群)に分けられています。これら20のグループの受容体には、PAMPsの認識に働くもの以外にも、植物ホルモンを認識するものや、発生・成長、器官形成を制御するものなど、さまざまな働きをするものがあります。植物ホルモンの認識や、発生・成長に関わるLRR-RLKsの中には、免疫型LRR-RLPsと同様にリガンド認識後にBAK1と結合して活性化するものがあります。しかし、免疫型LRR-RLPsと発生・成長型LRR-RLKsとの間に、どのような関連があるのか、なぜ同じ共受容体を必要とするのかは不明でした。
研究手法と成果
国際共同研究チームは一般に公開されている350種の植物ゲノム情報から、RLKをコードする約17万個の遺伝子を、RLPをコードする約4万個の遺伝子をそれぞれ抽出し、比較解析して受容体の進化を調べました。抽出した受容体の種類を調べたところ、LRR-RLKsおよびLRR-RLPsは植物の受容体の中で最も数が多く、RLKの約半数がLRR-RLKsであり、RLPの約7割がLRR-RLPsでした。
最近の研究から、免疫型LRR-RLPsは細胞外のLRRから突き出したアイランド領域(ID)[12]を持っており、IDがPAMPsの認識に重要な役割を果たすことが分かってきました。また、植物ホルモンを認識するLRR-RLKsや、発生・成長、器官形成を制御するLRR-RLKsの中にも免疫型LRR-RLPsと同様にIDを持つものが存在することが知られていました。そこで、免疫型LRR-RLPsとIDを持つLRR-RLKsとの関連を明らかにするため、LRR-RLPsおよびLRR-RLKsのうちIDを持つものを抽出し、LRRのどこにIDが存在するかを調べました。すると、IDを持つLRR-RLPsのほとんど全てがLRRの繰り返し配列の下から4番目と5番目の間にIDを持っていました。また、同じ場所にIDを持つLRR-RLKsを探索したところ、ほぼ全てのLRR-RLKsが、発生・成長を制御するLRR-RLKsグループXb(LRR-RLK-Xbs[13])に属していました。
次に、免疫型LRR-RLPsとLRR-RLK-Xbsの配列を比較したところ、IDの下にある四つのLRR領域の配列が酷似していました。これによって、この二つの受容体ファミリーは共通の祖先から派生したものであることが分かりました。さらに詳細に解析すると、この四つのLRR領域には、共受容体であるBAK1と結合するために必要なアミノ酸配列が存在していました。BAK1はリガンドが結合したLRR-RLKsおよびLRR-RLPsに結合する性質を持っています。よって、免疫型LRR-RLPsと発生・成長型LRR-RLK-Xbsは、祖先型の受容体からこの四つのLRR領域を受け継ぐことにより、リガンド依存的なBAK1との結合能力を保持していると考えられます(図1-1)。
一方で、免疫型LRR-RLPsと発生・成長型LRR-RLK-Xbsは、それぞれ独自の進化を遂げることで免疫と発生・成長という異なる反応を誘導する受容体へと分化したことが判明しました。これらの受容体は、IDおよびその上部のLRR領域の配列を多様化させて、PAMPsや内生ホルモンなどさまざまなリガンドを認識する能力を獲得したと考えられます(図1-2)。さらに、免疫型LRR-RLPsは細胞膜貫通領域[14]とその周辺領域に共受容体SOBIR1と結合するために必要なアミノ酸配列を獲得しました(図1-3)。そして、免疫型LRR-RLPsはPAMPsを認識するとBAK1およびSOBIR1という二つの共受容体と結合することで、二つの共受容体が持つリン酸化領域を近づけ、互いに活性化させることで、下流の免疫反応を誘導すると考えられます。一方、発生・成長型LRR-RLK-Xbsのグループは、細胞質領域にリン酸化酵素領域を獲得しました。そして、発生・成長型LRR-RLK-Xbsは植物ホルモンを認識すると、BAK1と結合して、自身のリン酸化領域とBAK1のリン酸化領域を近づけ、互いに活性化させることで、特異的に発生・成長に関わる反応を誘導できるようになったと考えられます(図1-4)。
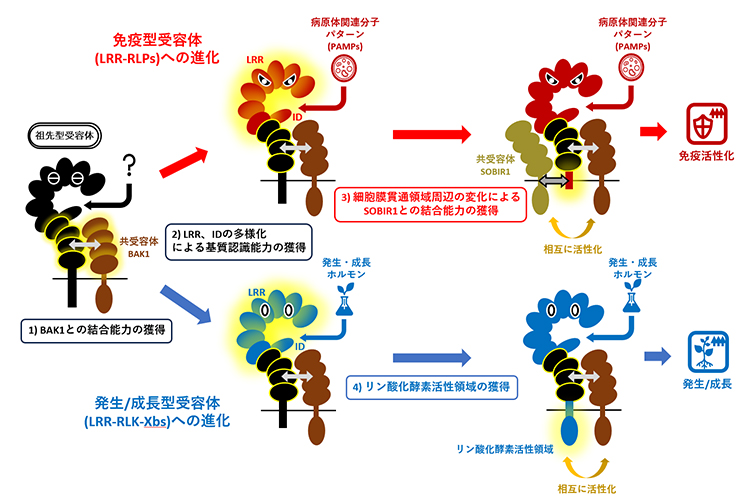
図1 免疫に働く受容体と発生・成長を担う受容体の進化モデル
免疫型LRR-RLPsおよび発生・成長型LRR-RLK-Xbsは共通の祖先からBAK1結合能力を獲得し、それぞれに必要な機能モジュールを進化させることで異なる受容体として分化した。
以上のように、免疫型LRR-RLPsおよび発生・成長型LRR-RLK-Xbsは、祖先型の受容体からBAK1と結合するために必要な領域を受け継ぎました。さらにSOBIR1との結合領域や、細胞質のリン酸化酵素領域など、機能的なモジュールを独自に獲得することで、別の受容体へと進化したと考えられます。
免疫型LRR-RLPsと発生・成長型LRR-RLK-Xbsの間で、これらのモジュールを入れ替えても機能することを確認しました。例えば、免疫型LRR-RLPsに発生・成長型LRR-RLK-Xbsのリン酸化酵素領域を結合させたキメラ受容体を作成したところ、キメラ受容体はPAMPsを認識して、免疫反応と発生・成長を制御する反応の両方を誘導しました(図2)。このことは、機能的モジュールを入れ替えることで新たな機能を持つ受容体を人工的に作成できることを示しています。
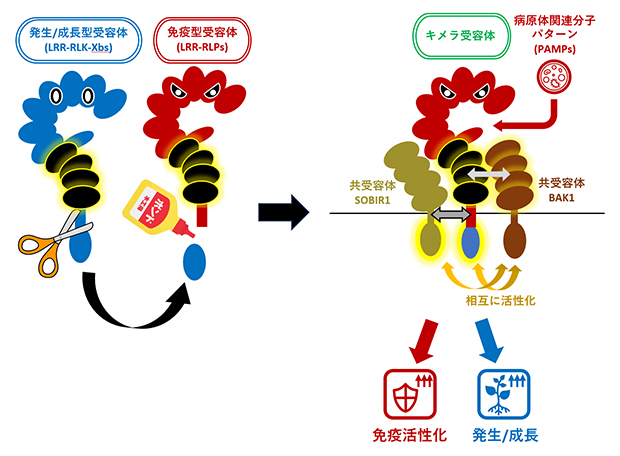
図2 免疫反応も発生・成長に関わる反応も誘導できるキメラ受容体
発生・成長型LRR-RLK-Xbsのリン酸化酵素領域を免疫型LRR-RLPsに結合したキメラ受容体はPAMPsを認識すると免疫反応と発生・成長制御の両方を誘導する。
今後の期待
植物免疫を理解する上で重要な疑問の一つであった植物免疫の起源について、本研究は分子レベルでの解答を与えることができました。さらに免疫型LRR-RLPsと発生・成長型LRR-RLK-Xbsの進化と、それぞれが進化させた機能的モジュールが明らかになりました。これにより、植物のゲノム情報から、免疫受容体として働く遺伝子と発生・成長に関わる遺伝子を簡便かつ正確に予測することができるようになりました。現在はこれらの情報を駆使して、さまざまな植物から免疫受容体を単離して解析しています。そして、これらの免疫受容体を活用することで、耐病性作物作出などへの応用に取り組んでいきます。
本研究成果は、国際連合が定めた17項目の「持続可能な開発目標(SDGs)[15]」のうち、「2.飢餓をゼロに」「15.陸の豊かさも守ろう」に貢献するものです。
補足説明
- 1.受容体
細胞外からやってくるさまざまなリガンド(神経伝達物質、ホルモン、病原菌など)を選択的に受容するタンパク質。細胞膜に存在するものや、細胞質あるいは核内に存在するものもある。 - 2.ロイシンリッチリピート(LRR)
疎水性のアミノ酸であるロイシンを多く含む20~30残基のアミノ酸の繰り返し配列で構成され、その構造はタンパク質や低分子などの認識に重要な役割を果たす。受容体タンパク質のリガンド認識部位として使われることが多い。典型的な受容体はLRRの繰り返しを20個以上持ちコイル状の構造をとる。LRRはLeucine-rich repeatの略。 - 3.ロイシンリッチリピート受容体型リン酸化酵素(LRR-RLKs)
LRR領域、細胞膜貫通領域、リン酸化酵素活性を持つ領域から構成される受容体で、植物のさまざまな情報伝達において重要な役割を果たすタンパク質ファミリー。LRR領域でリガンドを認識し、タンパク質のリン酸化反応を介して細胞内へと情報を伝える。RLKはReceptor-Like Kinaseの略。 - 4.ロイシンリッチリピート受容体様タンパク質(LRR-RLPs)
植物の細胞膜に存在する受容体ファミリー。LRR領域と細胞膜貫通領域から成る。LRR領域でリガンドを認識するが、細胞質側に機能的領域を持たないため、リン酸化酵素活性を持つ共受容体の助けを借りて細胞内へ情報を伝達する。RLPはreceptor-like proteinの略。 - 5.モジュール
タンパク質のモジュールとは、タンパク質内にあり特定の構造や機能を持つ領域のこと。例えば、他のタンパク質との結合を担う領域、酵素活性を有する領域などがこれに当たる。多くのモジュールを持つタンパク質では、それぞれのモジュールの機能が統合されてタンパク質全体の機能が発揮される。 - 6.リガンド
受容体に認識されるタンパク質や低分子。 - 7.リン酸化酵素領域
リン酸化酵素はタンパク質にリン酸基を付加する活性を持つ。このリン酸化は、基質タンパク質の構造や活性化などに影響を及ぼす。リン酸化酵素の多くは細胞内の情報伝達において重要な役割を果たす。細胞膜局在型のRLKは細胞質側にリン酸化酵素領域を持つ。これにより、RLKは外部からの刺激を認識するとリン酸化酵素反応により細胞質の因子へと情報を流す。 - 8.病原体関連分子パターン(PAMPs)
病原体に普遍的に存在し、宿主の免疫系によって認識される分子構造のこと。細菌の鞭毛タンパク質、糸状菌の細胞壁分解産物であるキチンなどが含まれる。PAMPsはPathogen-associated molecular patternsの略。 - 9.共受容体
受容体と結合・協調してリガンドの認識を担ったり、受容体の活性化や下流因子の活性化に寄与したりするタンパク質。 - 10.SOBIR1
LRR-RLKsの一つで、免疫型LRR-RLPsに結合する共受容体として働き、免疫型LRR-RLPsからの情報伝達に不可欠である。SOBIR1は恒常的に免疫型LRR-RLPsと結合しており、PAMPsの認識時にBAK1が免疫型LRR-RLPsと結合することで、SOBIR1とBAK1が互いに活性化し、協調して下流因子を活性化する。SOBIR1はSUPPRESSOR OF BAK1-interacting receptor-like kinaseの略。 - 11.BAK1
LRR-RLKsの一つで、植物ホルモン、ブラシノステロイド受容体と結合する共受容体として同定された。その後の研究により、BAK1は細胞膜型の免疫受容体や、細胞膜型のペプチドホルモン受容体の共受容体として働くことが明らかにされた。BAK1は受容体と共に基質の認識を担うとともに、受容体の活性化や下流因子の活性化に重要な役割を果たす。BAK1はBRI1 Associated receptor Kinase 1の略。 - 12.アイランド領域(ID)
LRR領域に存在する挿入領域。LRRの繰り返し配列とは異なるアミノ酸配列を持ち、PAMPsなどの基質との結合に寄与する。IDはIsland domainの略。 - 13.LRR-RLK-Xbs
LRR-RLKのグループの一つ。植物の成長、細胞分裂と増殖、種子の発芽などに関わる植物ホルモンであるブラシノステロイドの受容体BRI1や、細胞の増殖や分化促進を誘導するホルモンであるファイトスルフォカインの受容体であるPSKR1/2などが含まれる。 - 14.細胞膜貫通領域
細胞膜に存在するタンパク質のうち、膜貫通型タンパク質が持つ、細胞膜を貫通する領域。 - 15.持続可能な開発目標(SDGs)
2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された国際目標。持続可能な世界を実現するための17のゴールから構成され、地球上の誰一人として取り残さないことを誓っている。SDGsは発展途上国のみならず、先進国自身が取り組むユニバーサル(普遍的)なものであり、日本としても積極的に推進している。
国際共同研究チーム
理化学研究所 環境資源科学研究センター 植物免疫研究グループ
グループディレクター 白須 賢(シラス・ケン)
(環境資源科学研究センター 副センター長)
基礎科学特別研究員 ブルーノ・ポクマン・ゴウ(Bruno Pok Man Ngou)
専任研究員 門田 康弘(カドタ・ヤスヒロ)
Marc W Schmid 社(スイス)
研究員 ミシェル・ウィラー(Michele Wyler)
研究員 マーク・シュミッド(Marc W Schmid)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業基盤研究(A)「アポプラストにおける植物-病原体相互作用(研究代表者:白須賢)」、同基盤研究(B)「植物による線虫認識機構の解明(研究代表者:門田康弘)」、同特別研究員奨励費「害虫抵抗性を付与する植物免疫受容体の同定(研究代表者:白須賢)」、同挑戦的研究(萌芽)「アグロバクテリウムのステルス化による植物の形質転換効率の向上(研究代表者:門田康弘)」、文部科学省学術変革領域研究(A)「不均一環境変動に対する植物のレジリエンスを支える多層的情報統御の分子機構(研究領域代表者:松下智直)」の計画研究「不均一土壌環境に応答した寄生植物の感染統御機構(研究代表者:吉田聡子)、二国間交流事業共同研究「植物と寄生線虫の生物間コミュニケーションの分子機構解明(研究代表者:門田康弘)」による助成を受けて行われました。
原論文情報
- Bruno Pok Man Ngou, Michele Wyler, Marc W Schmid, Yasuhiro Kadota, Ken Shirasu, "Evolutionary Trajectory of Pattern Recognition Receptors in Plants", Nature Communications, 10.1038/s41467-023-44408-3
発表者
理化学研究所
環境資源科学研究センター 植物免疫研究グループ
グループディレクター 白須 賢(シラス・ケン)
(環境資源科学研究センター 副センター長)
基礎科学特別研究員 ブルーノ・ポクマン・ゴウ(Bruno Pok Man Ngou)
専任研究員 門田 康弘(カドタ・ヤスヒロ)
 門田 康弘、白須 賢、ブルーノ・ポクマン・ゴウ
門田 康弘、白須 賢、ブルーノ・ポクマン・ゴウ
報道担当
理化学研究所 広報室 報道担当
お問い合わせフォーム