2009年9月22日
独立行政法人 理化学研究所
独立行政法人科学技術振興機構
国立大学法人東京大学
劣悪環境に応答する植物ホルモン「アブシジン酸」の応答経路を解明
-植物の環境ストレス耐性の制御機構が明らかに-
ポイント
- タンパク質リン酸化・脱リン酸化酵素の複合体がアブシジン酸のON/OFFスイッチ
- 「アブシジン酸受容体→脱リン酸化酵素→リン酸化酵素」の単純な情報伝達経路が存在
- 干ばつや塩害、冷害など劣悪環境耐性作物の開発や種子品質の制御が容易に
要旨
独立行政法人理化学研究所(野依良治理事長)、独立行政法人科学技術振興機構(以下JST、北澤宏一理事長)と国立大学法人東京大学(濱田純一総長)は、劣悪環境への応答にかかわっている植物ホルモン「アブシジン酸(ABA)※1」のシグナル伝達経路※2を世界で初めて解明しました。これは、理研植物科学研究センター機能開発研究チーム(篠崎一雄センター長兼チームリーダー)の梅澤泰史研究員、基幹研究所中野生体膜研究室の平山隆志専任研究員(横浜市立大学客員教授)らと、慶應義塾大学の石濱泰准教授ら、横浜市立大学生命ナノシステム科学研究科および東京大学農学生命科学研究科の篠崎和子教授らによる共同研究の成果です。
植物は、根を下ろした場所から移動することができないので、土壌・水・日照などの生育環境が悪化しても、それに適応するしか生存の道はありません。そのため植物は、独自の適応能力を備えており、ABAはその中心的な調節物質であることが分かっていました。しかし、ABAの情報がどのように細胞内を伝わり、必要な応答が引き起こされるのかはよく分かっておらず、長年の課題となっていました。研究グループは、細胞内シグナル伝達の中で、特にタンパク質のリン酸化・脱リン酸化反応に着目して研究を行い、タンパク質脱リン酸化酵素の1つ「PP2C(Protein Phosphatase 2C)※3」が、タンパク質リン酸化酵素の1つ「SnRK2 (SNF1-related protein kinase 2)※4」と相互作用し、ABAシグナル伝達経路が調節されていることを世界で初めて明らかにしました。また、最近報告されたABAの細胞内受容体の一群である「RCAR/PYR※5」との関係も調べたところ、ABAが受容体に作用するところから環境ストレス耐性遺伝子が発現するところまでのシグナル伝達経路を確定しました。
ABAは、植物の耐乾燥性、耐塩性、耐冷性などの環境ストレス耐性を制御するだけでなく、病虫害への抵抗性などにも関与しています。さらに、種子成熟や発芽、成長、老化など植物のあらゆる生育段階に影響を持つ極めて重要なホルモンです。今回、ABAのシグナル伝達経路を解明したことで、ABAの作用を人為的に制御するための基盤を築き上げることができました。将来的には、干ばつ・塩害・冷害などに耐性を持つ作物の開発、種子の安定性向上や穂発芽※6の抑制など、さまざまな応用が期待できます。
本研究に用いた分析手法は、JST戦略的創造研究推進事業 個人型研究(さきがけ)の研究領域「構造機能と計測分析」(研究総括:兵庫県立大学 寺部茂名誉教授)における研究課題「オミクス解析用超微小エレクトロスプレー法の開発」(研究者:石濱 泰)で開発されたものです。本研究成果は、米国科学アカデミー紀要『Proceedings of the National Academy of Sciences of the USA』オンライン版に9月21日の週に掲載されます。
背景
昨今の地球規模での温暖化や、それに伴う天候不順、異常気象などにより、農業環境の悪化や作物収量の減少が懸念されています。一方で、人口増加による食料供給の不足に加え、代替エネルギーとしての需要増加により、作物収量増産の要求は急速に高まっています。この問題を打開する方策の1つとして期待されているのが、劣悪環境に耐性を持つ有用作物の開発です。現在、作物の収量減少の原因の半分が、干ばつ、低温、塩害といった、環境条件の変化によるものといわれており、作物のこれらの環境耐性能を引き上げることで、大幅な改善が見込まれます。
植物の環境耐性能は、主に植物ホルモンの1つであるアブシジン酸(ABA)によって制御されます(図1)。ABAは、乾燥などのストレスによって合成量が増加し、植物のさまざまなストレス応答を誘導します。例えば、ABAには葉の気孔を閉鎖する働きがあり、乾燥した際に蒸散による水損失を抑え、さまざまな遺伝子発現を誘導して耐性レベルを引き上げます。従って、ABAの作用機構を明らかにすることができると植物の環境応答の主要なメカニズムの理解につながり、耐性作物開発のための基盤となると期待できます。
ABAが植物細胞に作用するためには、ABAの信号を細胞内に伝えるシグナル伝達経路が必要です。このシグナル伝達では、経路上でシグナルのON/OFFを切り替える仕組みが重要となります。研究グループは長年この研究に取り組んでおり、それぞれ異なるアプローチで、タンパク質脱リン酸化酵素であるPP2Cや、リン酸化酵素であるSnRK2が、ABAのシグナル伝達にかかわることを明らかにしてきました。タンパク質のリン酸化は、生体シグナルの主要な伝達手段で、多くの生命現象のON/OFFを制御しています。当然、ABAシグナルについても同様の役割が予想されましたが、その具体的なメカニズムはなかなか解明されず、ABAシグナル伝達経路の全容は長い間不明なままでした。
研究手法と成果
SnRK2は、植物に特有のタンパク質リン酸化酵素の1種で、タンパク質をリン酸化して信号を伝達する機能を持ちますが、ABAによって活性化される特徴があります。SnRK2が破壊されたシロイヌナズナではABAに対する応答能力が失われることから、SnRK2はABAシグナルをONにする役割があると考えられます。しかし、SnRK2がどのような仕組みで活性化し、シグナルをONにするのかは不明でした。ABAシグナル伝達経路のもう1つの役者であるPP2Cは、タンパク質脱リン酸化酵素の1種であり、リン酸化されたタンパク質を元に戻す働きがあります。PP2Cは、SnRK2とは逆にABAシグナルをOFFにしますが、PP2Cの直接のターゲットはやはり不明なままでした。このため、ABAシグナル伝達の全容は、長い間未解決の問題として残されてきました。
研究グループは、PP2CとSnRK2が多くの面で対称的な性質(リン酸化/脱リン酸化、シグナルのON/OFF)を持つことから、これら2つが協調してABAシグナルを制御しているのではないかと予想しました。そこで、BiFC法※7などのさまざまな実験技術を駆使して、まずPP2CとSnRK2が植物細胞内で相互作用していることを証明しました(図2a)。次に、試験管内でPP2CとSnRK2を反応させ、PP2CがSnRK2を脱リン酸化して活性を抑える働きがあることを見いだしました(図2b)。さらに、SnRK2のリン酸化部位を独自のリン酸化プロテオミクス技術※8で詳しく解析したところ、SnRK2の活性化に伴って特定のアミノ酸残基がリン酸化を受け、PP2Cにより特異的にその残基が脱リン酸化されることが分かりました(図3)。つまり、PP2CによるSnRK2の不活性化が、ABAシグナルをOFFにする仕組みの実体であるということが判明しました。
最近、PP2Cの活性が、ABA受容体(RCAR/PYR)によって抑制されることが報告されました。この報告と今回の知見を合わせると、植物細胞内では[ABA受容体]→[PP2C]→[SnRK2]という複合体が形成され、ABAシグナル伝達経路の中枢として機能している可能性が考えられました。この予想を証明するために、研究グループはこれらの因子を再構成し、ABAシグナルのON/OFFを試験管内で再現することに成功しました。これにより、ABAが受容体に作用するとPP2Cが抑えられ、SnRK2が活性化する、というシンプルなシグナル伝達の実体が明らかとなりました(図4)。
研究グループが調査した結果、このABAシグナル伝達機構はコケ類から高等植物まで存在しており、それ以前の緑藻類には見いだされませんでした。このことから、植物の陸上化の進化過程において、このABAシグナル伝達機構が重要な役割を果たしたのではないかと推察されます。
今後の期待
環境問題や食料問題など、植物の育成、増産を促す技術開発が望まれています。本研究により、劣悪環境適応で主役を担うABAのシグナル伝達経路と、そこで機能する主要因子が明らかとなりました。これにより、ABAシグナルを人為的に制御する技術の開発が容易になると考えられます。具体的には、明らかにした主要因子を操作することでABAの作用を強めたり、逆に弱めたりすることが可能になると期待できます。ABAの作用を強めることで干ばつや塩害、冷害に強い作物ができることは、これまでの多くの研究が実証していますが、ABAは生育抑制作用も持つため実際の応用は困難でした。今回の成果は、ABAシグナルのON/OFFを人為的に操作することを可能にし、これまでの技術的な問題を克服できる可能性を秘めています。
ABAは古くから知られる五大植物ホルモン※9の1つで、図1に示したように、植物のさまざまな生理現象に作用しています。そのため、環境ストレス耐性以外にもさまざまな応用が期待されます。例えば、コムギやトウモロコシなどで問題となっている穂発芽を抑制し、穀物の品質を高めるといった応用も可能になります。逆に、ABAの作用を弱め作物の生育を促進させたり、種子発芽を誘導し生育時期を調整するなど、今回の成果の適用範囲はきわめて多岐にわたるのが特徴です。さらに、ABAのシグナル伝達経路とそこで機能する主要因子を明らかにした今回の成果は、ABAがかかわる生理現象の理解促進などの基礎科学面でも大いに貢献すると期待されます。
発表者
理化学研究所
植物科学研究センター 機能開発研究チーム
研究員 梅澤 泰史(うめざわ たいし)
Tel: 029-836-4359 / Fax: 029-836-9060
基幹研究所 中野生体膜研究室
専任研究員 平山 隆志(ひらやま たかし)
Tel: 045-508-7220 / Fax: 045-508-7363
応用生命化学専攻 植物分子生理学研究室
教授 篠崎 和子(しのざき かずこ)
Tel: 03-5841-8137 / Fax: 03-5841-8009
お問い合わせ先
横浜研究推進部 企画課Tel: 045-503-9117 / Fax: 045-503-9113
(JSTの事業に関すること)
独立行政法人科学技術振興機構
イノベーション推進本部 研究推進部(さきがけ担当)
原口 亮治(はらぐち りょうじ) Tel: 03-3512-3525 / Fax: 03-3222-2067
報道担当
独立行政法人理化学研究所 広報室 報道担当Tel:048-467-9272 / Fax:048-462-4715
独立行政法人科学技術振興機構 広報ポータル部
Tel: 03-5214-8404 / Fax: 03-5214-8432
補足説明
- 1.アブシジン酸(ABA)
植物ホルモンは、比較的低濃度で作用する植物生長調節物質であり、アブシジン酸(ABA)はその1つである。英語名abscisic acid、分子式C15H20O4で表される。ABAの代表的な生理作用として、気孔の閉鎖、乾燥耐性の獲得、種子成熟・休眠、落葉など器官脱離の促進などが挙げられる。また、最近では病害抵抗性にかかわっていることが示唆されており、ほかの植物ホルモンとの相互作用も盛んに研究されている。 - 2.シグナル伝達経路
生体内で、ある種のシグナルがほかのシグナルに変換され、連続して伝わる過程のことを指す。一般的にその担い手はタンパク質であることが多く、シグナル伝達因子と呼ばれる。シグナル伝達因子の中にはさまざまな種類があるが、タンパク質のリン酸化にかかわる因子はそのうちの1つである。この場合、シグナルがタンパク質のリン酸基という形に変換されて伝わることになる。 - 3.PP2C(Protein Phosphatase 2C)
タンパク質脱リン酸化酵素の1種で、植物のABA応答を負に制御する。1990年代に、ABAへの感受性が弱まったシロイヌナズナ変異体の原因遺伝子 ABI1として、最初に同定された。シロイヌナズナには9つの類似した遺伝子が存在する。 - 4.SnRK2(SNF1-related protein kinase 2)
タンパク質リン酸化酵素の1種で、シロイヌナズナには9つのファミリーが存在する。そのうち、SRK2D、E、Iの3つがABAによって強く活性化し、ABA応答を正に制御する。 - 5.RCAR/PYR
ABAの細胞内受容体の1群。シロイヌナズナには14個の類似した遺伝子が存在しており、ABAの存在下でPP2C活性を抑制することが報告されている。 - 6.穂発芽
植物の種子が穂に残っている状態で発芽してしまう現象のことを指す。商品価値が失われるため、トウモロコシやコムギなどで農業上の大きな問題となっている。穂発芽は種子の休眠が不完全なことに起因し、ABAの作用が深くかかわっている。 - 7.BiFC法
Bi-molecular fluorescent complementation法の略。黄色蛍光タンパクYFP(yellow fluorescent protein)をN末側半分(YFPN)とC末側半分(YFPC)に分割し、結合能を調べようとする2つのタンパク質のそれぞれに融合させた組み換えタンパク質を発現させ、蛍光を観察する方法。2つのタンパク質に結合能がある場合、YFPNとYFPCが近接しYFPが再構成され、蛍光が観察される。 - 8.独自のリン酸化プロテオミクス技術
慶應義塾大学の石濱泰准教授らが、科学技術振興機構さきがけ研究において開発したリン酸化タンパク質の解析技術。独自のリン酸化ペプチドの精製技術と、超微小エレクトロスプレー法を組み合わせた質量分析装置を用い、リン酸化タンパク質の分析の性能が、従来法と比較して、選択性が約100倍、同定効率が約10倍向上した。 - 9.五大植物ホルモン
オーキシン、サイトカイニン、アブシジン酸、ジベレリン、エチレンの5種類の植物ホルモンのこと。これらは、古くからその存在が知られており、植物に対する作用もよく調べられている。植物ホルモンには、ほかにジャスモン酸、ブラシノステロイドや、最近見つかったストリゴラクトンなどがある。
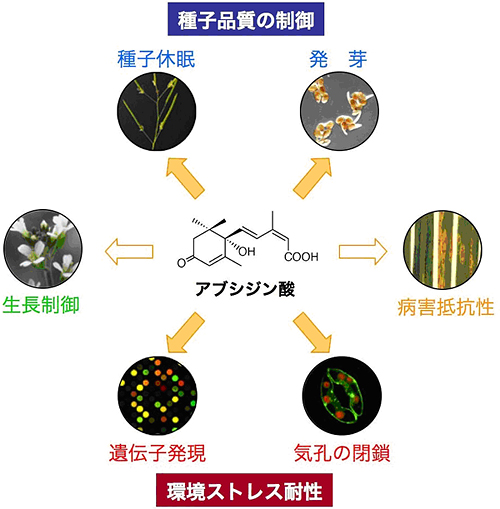
図1 アブシジン酸はさまざまな植物の機能をコントロールする重要な植物ホルモンである
植物が乾燥や高塩濃度などの劣悪環境に置かれると、植物の中でアブシジン酸(ABA)が増加し、遺伝子発現や気孔の閉鎖を促進することによって、劣悪環境に適応しようとする。また、ABAは種子の成熟過程でも増加し、種子休眠や発芽を制御する主要な植物ホルモンである。これだけでなく、ABAはさまざまな植物の生長制御にかかわっていることが知られており、さらに最近ではABAが病害抵抗性に関与していることが報告されている。
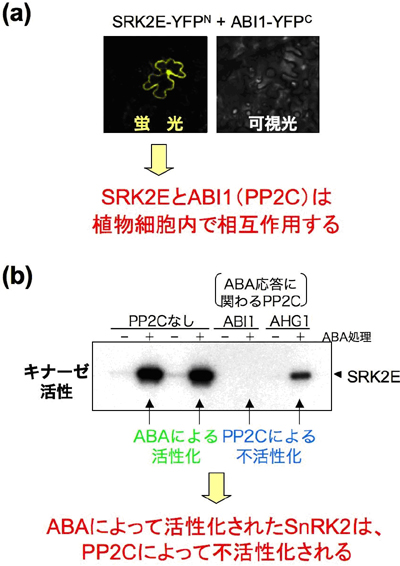
図2 PP2CとSnRK2は細胞内で相互作用する
(a) 植物細胞におけるABI1(PP2C)とSRK2E(SnRK2)の相互作用を、BiFC法によって調べた実験。ソラマメの葉の表皮において、それぞれ蛍光タンパク質の部分断片を融合したABI1とSRK2Eを発現させた。ABI1とSRK2Eが結合すると、蛍光タンパク質が再構成され、蛍光を発するようになる。
(b) ABA処理(+)または未処理(-)の培養細胞からSnRK2(SRK2E)を精製し、試験管内でPP2C(ABI1およびAHG1)と反応させ、リン酸化活性を観察した実験。黒いバンドがSRK2Eの活性を示しており、ABA処理を行なうとSRK2Eの活性化が観測される(PP2Cなし)。一方、この状態でPP2Cを反応させると、SRK2Eの活性が著しく減少した。これにより、PP2CがSnRK2を不活性化することが明らかとなった。
図3 PP2CによるSnRK2の不活性化と脱リン酸化の解析
図2と同様に調製したSRK2Eのリン酸化部位を、高速液体クロマトグラフィーおよび質量分析計により解析した結果。上の模式図はSnRK2の構造を示しており、今回検出したリン酸化部位は、プロテインキナーゼの活性化ループと呼ばれる重要な部分に相当する。下図は、黄色で示した部分に相当するリン酸化ペプチドのクロマトグラムである。解析の結果、このリン酸化ペプチドはABA処理によって増加し、PP2C (ABI1)反応により減少することを観測した。これにより、PP2CがSnRK2の活性化ループを直接脱リン酸化することが明らかとなった。
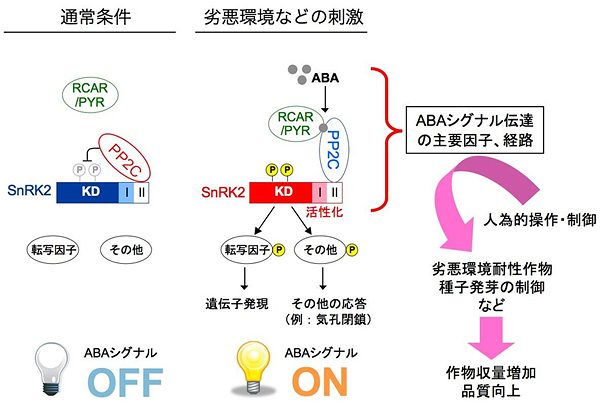
図4 本研究で明らかとなったABAシグナル伝達のメカニズム
通常の状態でABAが少ない時は、SnRK2に結合したPP2CがSnRK2を脱リン酸化し、活性化を抑えることでABAシグナルをOFFにしている。劣悪環境などの刺激を受けABAの蓄積量が増加すると、ABA受容体(RCAR/PYR)がPP2Cに結合し、PP2Cの活性を抑制する。その結果、SnRK2はPP2Cの抑制から解放され、転写因子などをリン酸化してこれらの因子の機能を調節し、ABAシグナルがONになってさまざまな応答が引き起こされる。本研究によって、ABAのシグナル伝達経路が単純化され、受容から遺伝子発現までをわずか4段階で説明できるようになった。今後は、これらのシグナル伝達経路を人為的に操作することで、劣悪環境耐性植物や種子発芽の制御など、農業的に有用な性質を付加した植物の開発に結びつくことが期待される。