2013年11月5日
理化学研究所
ゴルジ体内のタンパク質輸送を制御する分子機構の一端を解明
-新開発の顕微鏡システムによりRab GTPアーゼの転換機構を証明-
ポイント
- 新しい高感度共焦点顕微鏡システムを開発
- 複数のRab GTPアーゼ間の転換機構を解明
- ゴルジ体の成熟を担う分子機構の解明につながる
要旨
理化学研究所(理研、野依良治理事長)は、酵母を使い、細胞小器官のゴルジ体[1]でのタンパク質輸送を制御する分子機構の一端を解明しました。これは、理研光量子工学研究領域(緑川克美領域長)ライブセル分子イメージング研究チームの中野明彦チームリーダー、須田恭之研究員らの研究チームの成果です。
ヒトや酵母を含む真核生物の基本単位である細胞内には、細胞小器官と呼ばれるさまざまな構造体があり、膜構造による輸送経路(膜交通)により相互にタンパク質を輸送しています。その輸送を制御しているのが、Rab GTPアーゼ[2]というタンパク質です。ゴルジ体は、輸送されるタンパク質を糖鎖付加などにより修飾し、仕分けした後に適切な場所へ送り出す重要な機能を持っています。高等動植物のゴルジ体は、平らな袋(槽)が積み重なった構造をしており、シス槽、メディアル槽、トランス槽に分類され、それぞれの膜の成分や局在する酵素が異なっています。酵母のゴルジ体には4つ(Ypt1、Ypt6、Ypt31、Ypt32)のRab GTPアーゼが存在しますが、ゴルジ体の膜上でどのように時間的空間的に制御されているか、解明されていませんでした。
研究チームは、独自に開発した高感度共焦点顕微鏡システム(SCLIM)[3]を使用した生きた細胞のライブイメージングにより、酵母のゴルジ体に存在するRab GTPアーゼを可視化し、挙動を調べました。その結果、細胞小器官の1つであるエンドソームからゴルジ体へのタンパク質輸送を担うYpt6は、ゴルジ体が成熟するにつれて膜上から消失することが分かりました。さらに、Ypt6の挙動の制御には、Ypt6を不活性化させるタンパク質Gyp6がトランス槽に存在するYpt32と結合しトランス槽の膜上に局在する必要があることが明らかになりました。これらの結果は、近年提唱されているRab GAPカスケード[4]により一連のRab GTPアーゼ転換機構[5]が行われることを示しています。また、Rab GAPカスケードが正常に働かない変異株を作製したところ、Rab GAPカスケードがゴルジ体の機能、および成熟に少なからず寄与していることも明らかになりました。
今後、SCLIMによるライブイメージングを駆使し、ゴルジ体の成熟を司るメカニズムに迫ることで、ゴルジ体を中心とした細胞内膜交通の全容解明が期待できます。本研究成果は、米国科学アカデミー紀要『Proceedings of National Academy of Sciences of the United States of America』オンライン版に11月4日の週に掲載されます。
背景
全ての生物の構造上・機能上の基本単位である細胞の中には、細胞小器官と呼ばれるさまざまな構造体があります。ゴルジ体は細胞小器官の1つで、新たにつくられたタンパク質(積み荷タンパク質)を受け取り、糖鎖付加などの修飾を施し、仕分けして適切な場所へと正しく輸送しています。ヒトなどを含む高等動植物のゴルジ体は、平らな袋(槽)が積み重なった層板(スタック)構造をしており、積み荷タンパク質を受け取る側をシス槽、積み荷タンパク質が仕分けされ出て行く側をトランス槽、その中間をメディアル槽と呼びます。それぞれの槽は、膜の性質や局在する酵素群「レジデントタンパク質[6]」が異なります。ゴルジ体内の積み荷タンパク質の輸送については、過去に小胞輸送モデルと槽成熟モデルの2つが提唱されていました。
これまでに理研の研究チームは、出芽酵母を用いてゴルジ体内の輸送モデルの解明に取り組んできました。そして2006年、高速高感度のレーザー共焦点顕微鏡システムによるライブイメージングによって、シス槽が徐々にその性質をメディアル槽、トランス槽へと変えていく様子を捉えることに成功し、ゴルジ体内の輸送は槽が時間とともにその性質を変えていく槽成熟によることを示しました注)。しかし、現在でもゴルジ体内における積み荷タンパク質の輸送については論争が絶えず、ゴルジ体の成熟をつかさどる分子機構の解明が望まれています。
細胞内で行われている膜構造によるタンパク質の輸送経路(膜交通)を制御している分子の一つに「Rab GTPアーゼ」があります。Rab GTPアーゼは、GTP/GDP交換因子とGTPアーゼ活性化タンパク質によって活性化型、不活性型に交互に変換されます(図1)。活性化すると標的膜上でさまざまな膜交通を制御し、不活性化すると膜から細胞質中へと分散し、次の活性化に備えます。活性化型と不活性化型をサイクルすることからRab GTPアーゼは、分子スイッチと呼ばれています。酵母のゴルジ体にはRab GTPアーゼが4つ(Ypt1、Ypt6、Ypt31、Ypt32)存在し、ゴルジ体特有の膜交通を制御していることが知られていましたが、成熟に伴いその性質を時々刻々と変えていくゴルジ体の各槽においてどのように時間的・空間的に制御されているか、直接的には解明されていませんでした。そこで研究チームは、ゴルジ体に存在するRab GTPアーゼの挙動を、独自に開発した高感度共焦点顕微鏡システム(SCLIM)によってライブイメージングし、Rab GTPアーゼの制御機構の解明に取り組みました。
研究手法と成果
まず、研究チームは、3種のRab GTPアーゼとゴルジ体の各槽に局在するレジデントタンパク質とを異なる蛍光タンパク質で標識し、それぞれのRab GTPアーゼがどの槽に局在するか調べました。その結果、細胞小器官のエンドソームからゴルジ体への膜交通を制御するYpt6がトランス槽に局在する割合は低く、主にシス槽からメディアル槽に局在すること、多様な機能を持つYpt32は、Ypt6とは逆にトランス槽に局在することが分かりました。また、Ypt6の挙動をライブイメージングで詳細に解析したところ、ゴルジ体が成熟するにつれてYpt6はゴルジ体の各膜上から徐々に消失し、逆にYpt32は徐々に蓄積していました。この結果から、Ypt6とYpt32はゴルジ体の各膜上で相互排他的に存在することが分かりました(図2)。
2種のRab GTPアーゼの相互排他的な局在の制御には、近年提唱されている「Rab GAPカスケード」と呼ばれるRab GTPアーゼの転換制御機構が関与していることが考えられました。酵母にはRab GTPアーゼを不活性化型に変換するタンパク質(GTPアーゼ Activating Protein; GAP)が8種類存在します。そこで、それぞれの変異株を作製し、Ypt6とYpt32の局在を解析しました。その結果、GAPの1つであるGyp6を産生しないgyp6破壊株だけにYpt6とYpt32が同時に局在していました(図3)。
また、遺伝学的、生化学的解析により、Gyp6は活性化したYpt32が結合してゴルジ体膜上に局在し、そのGyp6がYpt6を不活性化し細胞質中に分散させていました。この分子機構により、Ypt6とYpt32は、ゴルジ体の膜上で相互排他的に存在することが分かりました。これは、ゴルジ体の成熟に伴い、Rab GAPカスケードによりYpt6が膜上から徐々に消失し、Ypt32が徐々に蓄積することで、それぞれが制御する膜交通が転換することを示しています(図4)。
一方で、Rab GTPアーゼがゴルジ体の各膜の性質を規定すると考えられています。そこで、Rab GAPカスケードのゴルジ体の槽成熟への寄与を調べました。具体的には、Rab GAPカスケードが機能せず、複数のRab GTPアーゼがゴルジ体の同じ膜上に共に存在する変異株を作製し、ゴルジ体の槽が成熟する時間を計測しました。その結果、変異株においても槽成熟は進むものの、野生株と比べて遅延が見られ、ゴルジ体の機能にも影響がありました。これらの結果はRab GAPカスケードがゴルジ体の槽成熟に少なからず寄与していることを示しています。
今後の期待
今回明らかになったタンパク質輸送に関わる分子機構は、ゴルジ体の槽成熟をつかさどる分子機構全体の一端です。本研究で明らかになったRab GTPアーゼをはじめとする膜交通を制御する分子群や、ゴルジ体の膜の組成などさまざまな因子が協調して成し遂げられていると考えられます。ゴルジ体は細胞内膜交通において、輸送すべき積み荷タンパク質が正確に運ばれるために行き先ラベルを正確に貼り、仕分け分別する配送センターとしての機能を果たしています。このような分子機構をひとつひとつ解明することでゴルジ体の機能の全容が明らかになることが期待できます。
ゴルジ体における輸送システムの破綻は高等生物では細胞、組織そして生態の恒常性維持に深刻な影響を及ぼします。真核生物のモデルとして長年研究が続けられる酵母を使って細胞の基本的なメカニズムを理解することが、ヒトや植物をはじめとする多細胞生物の生体の理解へとつながります。
本研究で開発、使用した顕微鏡システムは超解像技術の中でもライブイメージングに適しており、マルチカラーによるイメージングも可能になりつつあります。これらの技術を駆使することにより、今までに明らかにできなかった謎を解くことができると確信しています。
原論文情報
- Yasuyuki Suda, Kazuo Kurokawa, Ryogo Hirata and Akihiko Nakano.
"Rab GAP cascade regulates dynamics of Ypt6 in the Golgi traffic"
Proceedings of the National Academy of Sciences of the United States of America,2013,doi: 10.1073/pnas.1308627110
発表者
理化学研究所
光量子工学研究領域 エクストリームフォトニクス研究グループ ライブセル分子イメージング研究チーム
研究員 須田 恭之 (すだ やすゆき)
チームリーダー 中野 明彦 (なかの あきひこ)
お問い合わせ先
光量子工学研究推進室 広報担当
Tel: 048-467-9258 / Fax: 048-465-8048
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.ゴルジ体
1898年にイタリアの神経科学者カミロ ゴルジにより発見された真核生物に存在する細胞小器官。多くの生物では平らな袋状の槽が積み重なった層板(スタック)構造を示すが、出芽酵母ではスタック構造を示さず、各々の槽が細胞内に散在している。 - 2.Rab GTPアーゼ
Rasスーパーファミリーに属する低分子量GTPアーゼ(GTP結合タンパク質)。GTPに結合した活性化型とGDPに結合した不活性化型とをサイクルすることから分子スイッチと呼ばれ、活性化型は標的膜上でエフェクター分子と結合することにより、さまざまな細胞内の膜交通を制御している。 - 3.高感度共焦点顕微鏡システム(SCLIM)
SCLIMは、 Super-resolution Confocal Live Imaging Microscopyの略。ニポウディスク方式共焦点スキャナー、高性能のダイクロイックミラー、フィルターシステムによる分光器、冷却イメージインテンシファイアー(電子増倍管)と複数の高感度CCDカメラシステムから構成され、複数蛍光の同時取得と高S/N比の蛍光画像取得とを可能にした独自に開発した蛍光顕微鏡システム。 - 4.Rab GAPカスケード
活性化型のRab GTPアーゼがエフェクターとして他のRab GTPアーゼのGAPを結合し、そのRab GTPアーゼを不活性化型に変換する一連の機構。 - 5.Rab GTPアーゼ転換機構
同一の膜上において、機能するRab GTPアーゼが転換する機構。その分子機構としては、活性化型のRab GTPアーゼがエフェクターとして他のRab GTPアーゼのGEFを結合し活性化型に変換するRab GEFカスケードと、Rab GAPカスケードにより行われると考えられている。Rab GTPアーゼとそのエフェクタータンパク質は局在すべき標的膜が決まっており、Rab GTPアーゼ転換機構が膜の性質を規定している可能性も示唆されている。 - 6.レジデントタンパク質
ゴルジ体の各槽に特異的に定常的に局在するタンパク質で糖鎖修飾酵素やSNAREタンパク質がある。
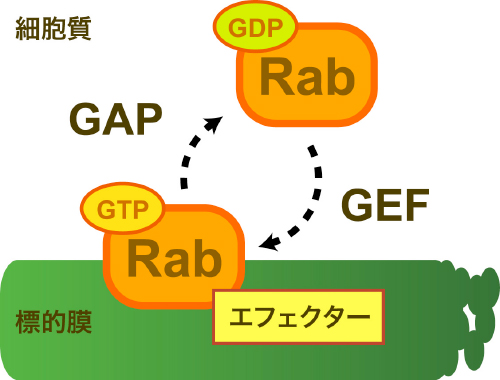
図1 Rab GTPアーゼの活性化サイクルとエフェクター
GTP/GDP交換因子であるGEFにより、Rab GTPアーゼは活性化型(GTP結合型)に変換する。活性化型は、標的膜上でエフェクタータンパク質と結合し、さまざまな膜交通を制御する。その後、GTPアーゼ活性化タンパク質であるGAPにより不活性化型(GDP結合型)に変換し、細胞質中に分散する。
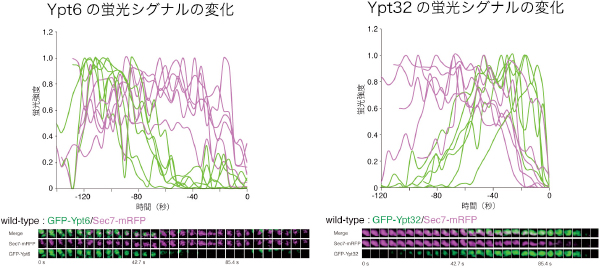
図2 野生株のゴルジ体におけるYpt6とYpt32の挙動と蛍光シグナルの変化
ゴルジ体トランス槽のマーカーSec7-mRFP(マゼンタ)におけるGFP-Ypt6もしくはGFP-YPt32(緑)の挙動(下図モンタージュ写真)。上図のグラフは独立した槽における蛍光シグナルの変化を示している。ライブイメージングの結果、ゴルジ体が成熟するにつれてYpt6はゴルジ体の各膜上から徐々に消失し、逆にYpt32は徐々に蓄積した。
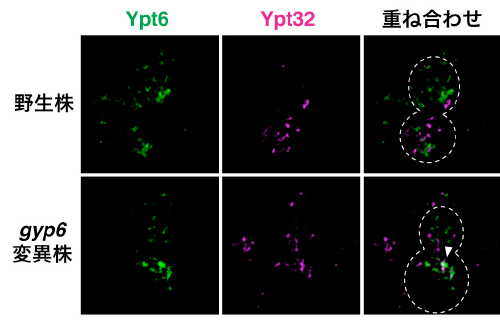
図3 野生株とgyp6変異株におけるYpt6とYpt32の局在
野生株とgyp6変異株におけるYpt6(緑)とYpt32(マゼンタ)の局在。野生株でそれぞれの蛍光シグナルは共局在しないが、gyp6変異株では矢じりで示すように共局在が見られた。点線は酵母細胞の形を示す。
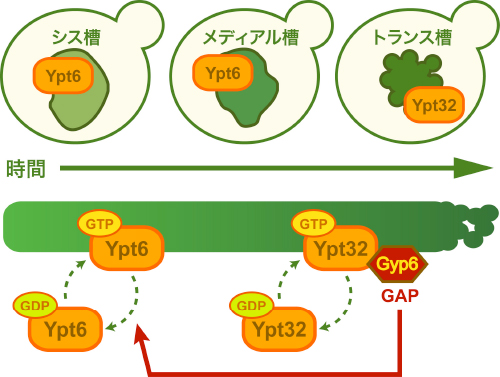
図4 Rab GAPカスケードによるYpt6からYpt32への転換
Gyp6は活性化型Ypt32と結合してゴルジ体膜上に局在する。そのGyp6がYpt6を不活性型に変換し、膜上から細胞質中に分散させる。図の上はゴルジ体の成熟と同調したYpt6からYpt32への転換を示す。