ポイント
- 気管が過剰に伸長するショウジョウバエ変異体を解析
- 気管上皮細胞の拡張力と細胞外マトリクスの弾性力が拮抗し、気管の長さを制御
- シンプルな物理モデルを構築して、同定したメカニズムのシミュレーションに成功
要旨
理化学研究所(理研、野依良治理事長)は、ショウジョウバエの胚において、細胞外マトリクス(基質)[1]の弾性力が気管の形態形成を制御していることを明らかにしました。そして、管の長さと形状の安定性は気管上皮細胞[2]の拡張力が細胞外マトリクスの弾性力と拮抗することで決まるという新しい仕組みを見いだしました。これは、発生・再生科学総合研究センター(竹市雅俊センター長)形態形成シグナル研究グループの董波(ボ・ドン)国際特別研究員(現 研究員)、林茂生グループディレクターらと、フランス キュリー研究所のEdouard Hannezo(エデュアルド・ハネッツォ)氏との国際共同研究グループの成果です。
生物の体内では、血管、呼吸管など管状組織のネットワークが縦横に張り巡らされ、血液や空気などの物質循環が行われています。物質がスムーズに循環するためには、部位ごとに管の太さが均一にそろっている必要があります。成長期のヒトの血管系では拍動の物理的な刺激に応じて血管内の細胞の再配置・増殖が起こり、血流が最適化されるように管の形状が変化します。しかし、胚発生においては血液循環が始まる前にすでに血管はある程度適正な形状を獲得しています。これまで、この循環に非依存的な管構造の適正化の仕組みはよく分かっていませんでした。過去の研究から、ショウジョウバエ胚の気管の内側(管腔)にはタンパク質とキチン質[3]を主成分とした細胞外マトリクスが蓄積し、気管の太さと長さという2つの形状特性を制御していることが知られていました。しかし、長さの制御において、細胞外マトリクスの果たす役割の詳細については未解明のままでした。
国際共同研究グループは、気管に蓄積する細胞外マトリクスの性状を詳細に解析したところ、粘り気がある液体成分と弾性体成分で構成されていることを見いだしました。これらの成分を損なう条件や、気管上皮細胞の管腔側の細胞膜(アピカル細胞膜[2])が過剰に拡大するショウジョウバエの変異体を解析し、物理モデルで検証しました。その結果、気管上皮細胞のアピカル細胞膜の拡張力と細胞外マトリクスの弾性力が拮抗することで力学的に安定し、適正な長さの気管ができることを見いだしました。
本成果は、細胞外マトリクスの形態形成に関わる新たな機能の発見であり、細胞が生物の管状組織を形作る細胞生物学的見地からも大変興味深い知見です。
本研究成果は、米国の科学雑誌『Cell Reports』(5月1日付け:日本時間5月2日)にオンライン掲載されます。
背景
ヒトを含めた大型の動物では、循環器系が栄養や酸素を体内の隅々に送り込むことで物資代謝の恒常性を保っています。管状組織のネットワークは動物の血管や気管などのように、体のサイズや各臓器の栄養要求度に合わせて柔軟に変化する可塑性を備えています。効率の良い循環を維持するためには、それぞれの個体や組織のサイズ、さらに管内を流れる物質の物理的性質などに合わせて、管の長さや太さを適切に調整する必要があります。また、管内を物質がスムーズに循環するには、部位ごとに管の太さが均一にそろっていることが不可欠です。脊椎動物の血管系では、心臓の拍動に合わせて血管に圧力がかかり、その物理的刺激が血管内皮細胞の再配置や増殖を促して、管の形状を最適化することが知られています。しかし、胚発生においては血管の太さや長さは心臓が動き始め血液が流れる前におおむね決まっていますし、気管ではそもそも拍動による刺激がなくても正しい形状をとることができます。このことから、管の形状を決める仕組みは発生過程のある段階で細胞や組織に備わっていると考えられます。
ショウジョウバエの気管は、各体節から陥入し、分岐・融合して一体のネットワークを形成します(図1)注1)。気管の内側(管腔)は当初は狭く、ほとんど閉じていますが、キチン質を主成分とした細胞外マトリクスが蓄積することで拡張します。その後、気管は孵(ふ)化後の一令幼虫の体長に合った長さにまで伸長したあとで管腔の細胞外マトリクスは細胞に吸収され、ガスに置き換わることで、孵化後の呼吸の準備が整います。また、管を形成する上皮細胞では、管腔に接する頂端側の細胞膜(アピカル細胞膜)と側方、基底側の細胞膜は異なった組成の膜タンパクと裏打ちのアクチン細胞骨格を持ち、それぞれ独自の経路により生産されています。
これまで多くの研究者により、気管が過剰に伸長するショウジョウバエの突然変異体が見いだされてきました。その原因遺伝子の解析から、気管の長さ制御にはアピカル細胞膜の合成、細胞接着分子の機能などによる複雑な経路が関与することが知られていました。さらに、キチン修飾酵素Serp[3]も気管の長さ制御に関わることから、細胞外マトリクスが気管伸長のブレーキとして働くことが示唆されてきました注2)。しかし、長さ制御の仕組みにおいて、細胞外マトリクスが具体的にどのようにして働くかについては不明のままでした。そこで国際共同研究グループは、細胞外マトリクスの物理的性質が長さ制御に関わる仕組みの探索に挑みました。
- 注1) 2013年1月14日プレスリリース
- 注2) 2013年1月16日プレスリリース
研究手法と成果
国際共同研究グループは、小胞輸送システムにおいて小胞形成時の細胞膜の陥入を制御する「ESCRTIII複合体[4]」の構成因子「Vps32[4]」に着目しました。Vps32は、遺伝子shrub[5]にコードされ、shrubに異常があるショウジョウバエ変異体は気管の過剰伸長を示しました(図2)。また、ESCRTIIIは膜タンパク質の輸送に重要であることが知られますが、shrub変異体を詳しく調べると、膜タンパク質である細胞接着分子の多くが細胞質内に蓄積する異常が見られました。特にアピカル細胞膜合成の促進因子「Crumbs[6]」は、通常は管腔に接する面(アピカル面)に蓄積しますが、変異体では細胞内小胞に蓄積しており、その蓄積量も増大しました(図3)。また、Crumbsは細胞質領域を通じてシグナルを発信しますが、shrub変異体では、異所的に局在したCrumbsがシグナルを過剰に発信するため、アピカル細胞膜合成が過剰になることが分かりました(図2)。
shrub変異体の気管は過剰に伸長する一方で、管腔の太さは正常胚と変わりませんでした。また、アピカル細胞膜が接する管腔は蛇行しますが、気管上皮細胞の基底膜の形状に大きな変化はありませんでした(図4)。これらのことから、shrub変異体では、アピカル細胞膜が長軸方向に過剰に伸長することで管自体が過剰伸長しますが、管腔の太さを広げる方向には変化しないことが明らかになりました。
管腔内の細胞外マトリクスには、キチン質の成分に加えて、高分子のタンパク質成分が存在します。その1つが、巨大膜結合タンパク質「Dumpy[7]」です。正常なショウジョウバエのDumpyタンパク質の性状について、GFP融合タンパク質を利用した光褪色後蛍光回復法(FRAP法[8])で調べたところ、Dumpyはいったん管腔内に放出されると安定な状態で存在し、ほとんど移動せずに気管の伸長に伴って引き延ばされることが分かりました(図5)。このことからDumpyは弾性を持ったゲル状態にあると考えられました。一方、低分子のGFP分子に対してもFRAP解析を行って管腔内の他の分子の状態を調べると、GFPはゆっくりと移動することから、Dumpyの他に粘性の高いゾル状態の成分も存在することが分かりました。
細胞外マトリクスで構成される昆虫の外骨格は、表皮細胞に密に結合しています。過去の研究から、Dumpyに糖鎖を付加する分子「EOGT」の変異体では、外骨格が表皮細胞のアピカル面からはがれることが知られています注3)。EOGT変異体では気管が過剰な伸長を示すことから、気管の長さ制御には、表皮細胞と細胞外マトリクスの結合が維持されることが必要であると考えられます。
これらの知見および考察をもとに、気管形成の様式をシミュレーションする物理モデルを構築しました。まず、弾性力を持つ円柱状の細胞外マトリクスを中心に据え、その周囲をアピカル細胞膜が包む管を想定します(このとき、単純化するため細胞の詳細や細胞個別の違いは無視します)。ここで、アピカル細胞膜に拡張力を与えると、細胞外マトリクスの弾性力が釣り合うところまで管が伸長しました。また、管の両端を固定した上で、アピカル細胞膜の拡張力が細胞外マトリクスの弾性力を上回るような条件を設定すると、気管が過剰に伸長して一定の波長で周期的に湾曲する様子が再現されました。このモデルにより、実際のshrub、dumpy、serpの変異体やそれらの組み合わせで起こる気管の形態変化をうまく説明することができました。
これらの結果から、気管の長さと形状の安定性は気管の上皮細胞の拡張力と細胞外マトリクスの弾性力が拮抗することで決まるという新しい仕組みが明らかになりました。
- 注3) Sakaidani, Y.et al.2011. O-linked-N-acetylglucosamine on extracellular protein domains mediates epithelial cell-matrix interactions. Nat Commun2, 583.
今後の期待
今回、ショウジョウバエの気管の長さと形状を決める仕組みにおいて、細胞膜と細胞外マトリクスの力学的相互作用が果たす新たな役割を見いだしました。細胞外マトリクスは、気管形成後も気管上皮細胞の働きで常に修飾を受け続けていることが分かっています。研究グループは、気管上皮細胞がキチン修飾酵素Serpを管腔からリサイクルして再分泌することで、気管の過剰伸長を防いでいることを報告しています。伸びる細胞が同時にブレーキをかけることで細胞外マトリクスの状態を更新しつつ、管の形を整えるという仕組みが明らかになりつつあります。
原論文情報
- Bo Dong, Edouard Hannezo, Shigeo Hayashi. "Balance between Apical Membrane Growth and Luminal Matrix Resistance Determines Epithelial Tubule Shape". Cell Reports,2014, http://dx.doi.org/10.1016/j.celrep.2014.03.066
発表者
理化学研究所
発生・再生科学総合研究センター 形態形成シグナル研究グループ
グループディレクター 林 茂生(はやし しげお)
お問い合わせ先
発生・再生科学総合研究センター 国際広報室
泉 奈都子(いずみ なつこ)
Tel: 078-306-3310 / Fax: 078-306-3090
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.細胞外マトリクス(基質)
細胞と細胞の間を満たし、生体組織を包み込む高分子の構造体。組織の支持体となるだけでなく、細胞外環境の情報を伝えることで細胞の増殖や分化を制御する。管状の組織では、管の内側(管腔)と外側で異なる細胞外基質が分布する。発生中のショウジョウバエ気管の管腔にはキチン質の多糖高分子やタンパク質からなる細胞外基質が満たされ、細胞表面を覆う外骨格(クチクラ)を形成するとともに、気管の太さと長さの制御を行うことが知られている。 - 2.上皮細胞、アピカル細胞膜
上皮細胞は組織を構築する基本的な細胞形態の1つで、主に表皮や臓器の粘膜、腺などを作る。細胞膜がアピカル(頂端側)とベーサル(基底側)の領域に分かれ、領域の境界部に細胞接着構造(アドヘレンスジャンクション)が鉢巻状に集積して、細胞同士を連結している。その結果、上皮細胞は共にアピカル面とベーサル面を持つシート状の構造をとり、条件によっては閉じた袋や管の形状をとる。
また、上皮細胞の細胞膜は、アピカル側(アピカル細胞膜)とベーサルおよび側方側とでタンパク質の組成が大きく異なり、それぞれ独自の物質輸送経路により生産される。 - 3.キチン質、キチン修飾酵素Serp
多糖高分子のキチンは、Nアセチルグルコサミンの多量体からなり、節足動物の外骨格の主成分となる。また、脱アセチル化反応により親水性の高いキトサンとなり、物理化学的性質を変化させる。ショウジョウバエ Serpentine( Serp)遺伝子の産物であるキチン修飾酵素Serpは、キチン−キトサン変換反応を触媒する酵素であると考えられている。この働きが低下すると気管は過剰に伸長することが知られている。 - 4.ESCRTIII複合体、Vps32
ESCRT複合体は0、I、II、Ⅲ、IVと順次細胞膜の細胞質側に結合して、細胞膜を陥入させて切り出す一連の反応を行う。ESCTRIII複合体はVps32などの7種類の分子が螺旋状に配列し、細胞膜をくびれ切る反応を媒介する。細胞質分裂質に伴う細胞膜の切断、小胞体内部への細胞膜の陥入反応、細胞膜に被覆されたウイルス粒子の放出など多様な反応に関わる。 - 5.shrub
ショウジョウバエVps32ホモログ(相同体)の遺伝子名。末梢神経の樹状突起は多数の分岐が幼虫の表皮を体内から広く森林状に覆う形状をとるが、 shrub変異体では樹状突起の分岐が縮退し、小さな茂み様に見える表現型から命名された。 - 6.Crumbs
ショウジョウバエで発見された細胞膜貫通分子。細胞内領域からアピカル細胞膜の形成を促進するシグナルが発信される。細胞外領域はアピカル細胞膜への集積とシグナル活性の調節を行う。変異体ではアピカル細胞膜が減少し、クチクラ表皮の形成が不全となる。変異体に見られる断片化したクチクラをパン屑に見立ててCrumbsと命名された。 - 7.Dumpy
ショウジョウバエで発見された遺伝子で変異体の翅が野生型に比べてずんぐりとした形状をとることから命名された。ショウジョウバエゲノム最大の遺伝子で、タンパク質は分子量250万を超える。合成されたDumpy分子は細胞外に放出され、C末端近くの細胞膜貫通領域で細胞に結合する。細胞外マトリクッスの構成分子で、変異体ではクチクラ構造の強度低下が認められる。 - 8.FRAP法
光褪色後蛍光回復法(Fluorescence Recovery After Photo bleaching)の略称で組織内の分子の移動、生産、分解などを調べる手法。組織や細胞が生きている状態で、GFPなどで蛍光標識された分子に局所的に強い吸収波長の光を与えて蛍光を退色させる。退色した領域での蛍光の回復速度とパターンを解析することで、分子の移動や生産に関する定量的な情報を得ることができる。
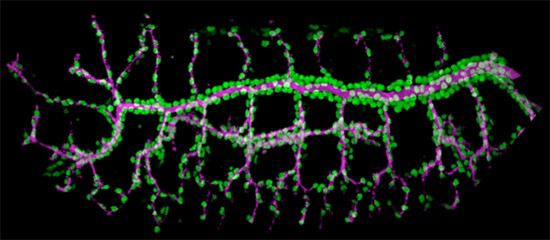
図1 ショウジョウバエ胚の気管系
胚発生後期にほぼ完成した気管系。気管細胞の核を緑で、管腔内の細胞外マトリックスをマゼンタで示す。

図2 shrub変異体では気管が過剰に伸長する
shrub変異体では気管の主要な管(DT)が正常胚(Control)に比べて過剰に伸長し、蛇行する。この表現型はcrumbs(crb)遺伝子を1コピー減らすと有意に回復するのでCrumbsタンパク質の過剰な活性化が関与すると考えられる。管腔にあるキチンの染色像(CBP)。スケールバー:50μm。
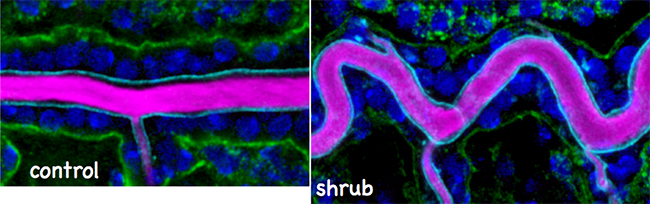
図3 shrub変異体での気管の形状
正常胚(Control)ではアピカル細胞膜(シアン)と基底膜(緑)がほぼ並行になっており細胞の高さが一定であることが分かる。一方、shrub変異体では過剰伸長したアピカル細胞膜が一定の管腔サイズを保ったまま蛇行する。基底膜は比較的まっすぐな形態を保つことから、場所によって細胞の位置や高さが著しく変わっていることが分かる。細胞核(青)の個数で判断できる細胞数は、両者に違いはない。
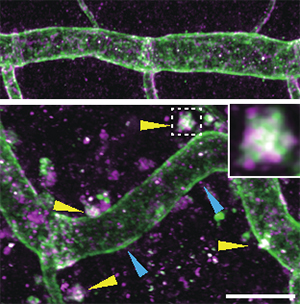
図4 shrub変異体ではCrumbsが小胞体に蓄積する
正常胚(上)ではCrumb(マゼンタ)とアピカル細胞膜マーカー(緑)はほとんどアピカル細胞膜に局在する。shrub変異体(下)では細胞質にマゼンタと緑のマーカーが混在して白く見える顕著な集合体が現れる(黄色矢印)。Crumbや膜タンパク質が異常に蓄積した小胞体だと考えられる。
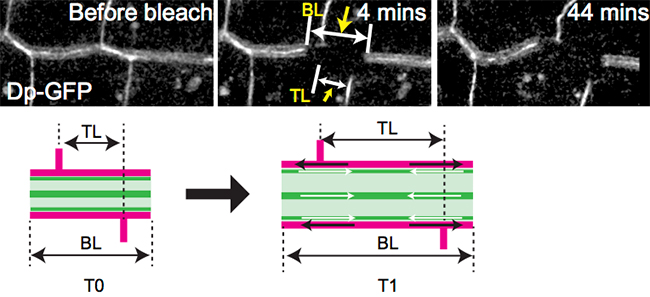
図5 Dumpyマトリクッスは伸長する
- (上段左)GFP標識されたDumpyタンパク質(Dp-GFP)の気管管腔での分布。
- (上段中央)光照射による蛍光退色4分後。BLは退色した細胞外マトリクッス部分の長さを、TLは上下への気管分岐を挟む気管の長さをあらわす。
- (上段右)退色44分後。退色したBL部分の蛍光はほとんど回復しておらず、むしろ拡大している。また、BLの拡大に伴ってTLも拡大している。このことから、Dp-GFP分子は管腔内でほとんど移動せず、また新たに合成される頻度も低いこと、さらに気管の伸長に伴ってDumpyタンパク質自体が引き延ばされていることが分かった。
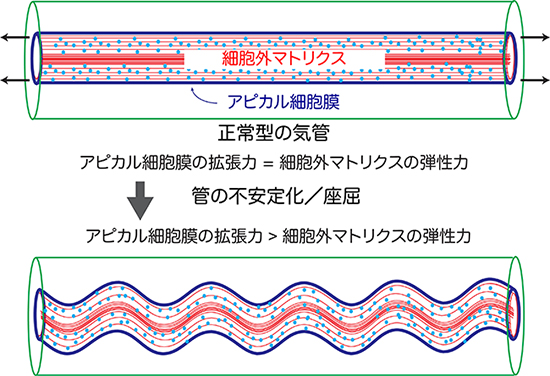
図6 気管伸張の物理モデル
- (上) 弾性を持った細胞外マトリクスを、シート状のアピカル細胞膜が包む。細胞の基底部側を緑で示す。アピカル細胞膜の拡張力が細胞外マトリクスの弾性力に釣り合うと、管は安定する。
- (下) アピカル細胞膜の拡張力が細胞外マトリクスの弾性力を上回ると管は伸長し、両端が固定された状態では周期的に湾曲する。