ポイント
- 覚醒は大脳皮質抑制性神経細胞を活性化し興奮性神経細胞の反応減衰を促進
- 前脳基底核から大脳皮質へのアセチルコリン投射系が抑制を強め覚醒効果を起こす
- 視覚情報処理の効率をあげる回路設計などへの応用が期待
要旨
理化学研究所(理研、野依良治理事長)は、覚醒や注意によって感覚が鋭敏になる脳の仕組みの解明に成功しました。覚醒状態では、脳の神経伝達物質であるアセチルコリン[1]の投射系が活性化し、大脳皮質の抑制性神経細胞(介在細胞)[2]の反応が増大することで、興奮性神経細胞(錐体細胞)[3]の反応がより速く減衰して、次にくる刺激に反応しやすくなることが分かりました。これは、理研脳科学総合研究センター(利根川進センター長)大脳皮質回路可塑性研究チームの木村塁研究員、サファリ・ミラ・シャハラム研究員、津本忠治チームリーダーらの研究チームによる成果です。
覚醒しているときや何かに注意をはらっているときに感覚が鋭敏になることは、良く知られており、心理学でもポピュラーな研究テーマです。視覚の場合、目から入った視覚情報は脳内で中継され、大脳後頭葉にある視覚野の神経細胞が反応して、最終的に認識されるとされています。つまり、どのように見えるかは、視覚野の神経細胞の「視覚刺激に対する反応」によって決まると考えられています。これまで、覚醒や注意によって視覚野の神経細胞の反応がどう変わるか研究されてきましたが、その仕組みは未解明のままでした。
大脳皮質の神経回路は、興奮性の錐体細胞と抑制性の介在細胞で構成されています。そこで、研究チームは、興奮性と抑制性の神経細胞を区別できる遺伝子改変ラットやマウスを用い、多数の神経細胞の活動を同時に記録可能な2光子励起カルシウムイメージング法[4]を使って大脳皮質視覚野の神経細胞の反応を詳細に調べました。その結果、動物が麻酔状態から覚醒状態に移ると、抑制性神経細胞の反応が増大し、興奮性神経細胞の活動がより速く減衰することを発見しました。また、覚醒に伴うこのような変化は、アセチルコリン投射系が抑制性神経細胞を活性化することによって生じることを明らかにしました。
今回の発見は、覚醒や注意をはらうことによって活性化される「感覚鋭敏化脳回路」とその動作の仕組みを明らかにしたもので、ロボットや、ウェアラブルコンピュータに用いる視覚情報処理のための回路設計などへの応用につながる可能性があります。
本研究成果は、米国の科学誌『Journal of Neuroscience』(7月23日号)に掲載されます。
背景
覚醒しているときや注意をはらっているときに感覚が鋭敏になることは、良く知られており、心理学でもポピュラーな研究テーマです。注意をはらっているときには素速く動くものやチカチカと点滅する視標を認知しやすくなることがその例です。感覚系のなかでも視覚に関しては、1981年にデイヴィッド・ヒューベルとトルステン・ウィーセルがノーベル生理学・医学賞を受賞して以来、多数の研究があり、大脳皮質視覚野の基本的な神経回路についての解明はかなり進んでいます。目から入った視覚情報は脳内で中継され、大脳後頭葉にある視覚野の神経細胞が反応して、最終的に認識されます。つまり、どのように見えるかは、視覚野の神経細胞の「視覚刺激に対する反応」によって決まると考えられています。しかし、覚醒や注意によって視覚野の神経細胞の反応がどう変わるか、その仕組みは未解明のままでした。
今回、研究チームはラットやマウスの大脳皮質視覚野を対象に、麻酔状態から覚醒状態に移ると神経細胞の反応がどのように変わるかを調べ、覚醒や注意によって感覚が鋭敏になる仕組みの解明に取り組みました。覚醒による脳機能の変化には、前脳基底核から大脳皮質に投射するアセチルコリン投射系(図1A)の関与が示唆されています。そこで覚醒に伴う視覚野の神経細胞の反応の変化にアセチルコリン投射系が関与しているかどうか、関与しているとすれば視覚野神経回路のどの部分にどのように関与するかを調べました。さらに主に2種類あるアセチルコリン受容体(ニコチン受容体とムスカリン受容体)のどちらがどのように関与しているかを詳細に調べ、アセチルコリンによって覚醒状態になる仕組みの全容解明を試みました。
研究手法と成果
研究チームは、多数の神経細胞の活動を同時に記録するため、少数の神経細胞しか記録できなかった微小電極法[5]に替え、2光子励起カルシウムイメージング法を用いました(図2)。また、活動した神経細胞が興奮性か抑制性かを同定できるように、抑制性神経細胞だけが緑色蛍光を発する遺伝子改変ラットを使用しました。実験の結果、ラットが麻酔状態から覚醒状態になると、抑制性神経細胞の視覚刺激に対する反応が増大すること、興奮性神経細胞の視覚刺激に対する反応がより速く減衰することを見いだしました(図3A)。
次に、前脳基底核から大脳皮質に投射するアセチルコリン投射系が、このような覚醒に伴う変化に関与しているかを調べました。麻酔下にあるマウスの前脳基底核を電気刺激して視覚反応の変化を調べたところ、覚醒時にみられたような抑制性神経細胞の反応増大と興奮性神経細胞の反応減衰の促進がみられました(図1B、図3B)。ただ、前脳基底核の電気刺激ではアセチルコリン投射系以外の細胞や神経線維も刺激している可能性を排除できません。そこで、光遺伝学[6]を用いて、アセチルコリンを含んだ神経細胞だけを青色光で活性化できるチャンネルロドプシン2遺伝子改変マウスを使ってアセチルコリン投射系の選択的活性化を試みました(図4)。その結果、抑制性神経細胞では視覚刺激による活動電位の発射頻度が増加すること、興奮性神経細胞では視覚刺激の後半に膜電位が抑制されることを見いだしました。
アセチルコリン受容体は大きく2種類(ニコチン受容体型とムスカリン受容体型)に分けられますが、どの受容体がこれらの変化に関与しているかを受容体拮抗薬を用いて脳スライス標本で調べました。その結果、大脳皮質第1層に存在する抑制性神経細胞では、主にニコチン受容体が機能していること、さらに2/3層に存在する抑制性神経細胞ではニコチン受容体とムスカリン受容体が共に働いていることが分かりました。これらの結果から、覚醒によるアセチルコリン投射系の活動によって活性化される神経回路と各受容体の作用部位が明らかになりました(図5)。
今回の成果により、覚醒や注意時にはアセチルコリン投射系が活性化することによって、まず抑制神経性細胞の反応が増大し、その結果、興奮性神経細胞の反応がより速く減衰して、次にくる刺激に反応しやすくなることが分かりました。
今後の期待
覚醒だけでなく、集中したりあるいはなにか強い動機をもったりして意欲的な行動をするときにも、私たちの感覚は鋭敏になります。今後は、覚醒に伴う神経回路機能の変化の研究に加え、注意集中や動機付けのような意欲に伴って感覚情報処理回路がどのように変化しその機能が向上するのか、さらにはその情報処理回路が感覚情報の変化に対応してどのように効率を変化させていくのかを明らかにしていく予定です。このような感覚神経回路機能の動的変化メカニズムを解明することで、例えば、ロボットの動作が対象や環境の変化に対応してその場にふさわしいようにする制御回路、あるいは身につけて持ち歩くことが可能なウェアラブルコンピュータにおいて、注意によって自動的に機能が変化する制御回路の設計などへの応用につながる可能性があります。
原論文情報
- Kimura Rui, Safari M-S, Mirnajafi-Zadeh J, Kimura Rie, Ebina T, Yanagawa Y, Sohya K, Tsumoto T "Curtailing effect of awakening on visual responses of cortical neurons by cholinergic activation of inhibitory circuits" Journal of Neuroscience, 2014,doi: 10.1523/JNEUROSCI.0863-14.2014
発表者
理化学研究所
脳科学総合研究センター 大脳皮質回路可塑性研究チーム
チームリーダー 津本 忠治(つもと ただはる)
お問い合わせ先
脳科学研究推進室
Tel: 048-467-9757 / Fax: 048-462-4914
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.アセチルコリン
神経伝達物質の一種で神経線維の末端から放出され、他の神経細胞に神経信号を伝える。その受容体は、イオンチャネル直結型のニコチン受容体と他のタンパク質を介して離れた部位にあるイオンチャネルの開閉に関与する代謝調節型のムスカリン受容体との2つに大別される。 - 2.抑制性神経細胞(介在細胞)
主にガンマアミノ酪酸 (GABA:γ-aminobutyric acid)を放出し、他の神経細胞の活動を抑制する神経細胞。軸索は通常短く、介在ニューロンとも呼ばれる。その数は大脳皮質視覚野では神経細胞全体の15~20%といわれている。 - 3.興奮性神経細胞(錐体細胞)
錐体細胞は、大脳皮質に多数(全神経細胞の80~85%)存在する神経細胞で、細胞体は錘体様の形状を示す。軸索末端からグルタミン酸を放出し他の神経細胞を興奮させることから興奮性神経細胞とも呼ばれている。通常、その軸索を遠くにある標的神経細胞へ伸ばす(投射する)。樹状突起はスパインと呼ばれる棘突起を豊富に持つ。 - 4.2光子励起カルシウムイメージング法
神経細胞の活動を可視化する方法の1つ。活動に応じて増減する細胞内カルシウムによって蛍光強度や波長が変化するカルシウム指示薬を細胞内に取り込ませ、神経細胞の活動を蛍光信号に変えて可視化する。従来の1光子励起顕微鏡は深部からの記録がぼけて鮮明な画像を得ることが困難であるが、深部からも空間分解能の良い画像の得られる2光子励起走査蛍光顕微鏡を使って神経細胞の活動を画像化する。 - 5.微小電極法
先端の直径が1μm~数μm以下の細い電極を脳に刺入し、直径数μm~20μm程度の小さな神経細胞が出す神経信号を細胞外や細胞内から記録する方法。 - 6.光遺伝学
遺伝子操作によって光で開閉するイオンチャンネルであるチャネルロドプシン2やハロロドプシンが特定の神経回路や神経細胞だけに発現するようにすると、その神経回路や神経細胞を光照射で活性化あるいは不活性化できる。この方法によって神経回路機能を解明しようとする研究手法を光遺伝学と呼ぶ。
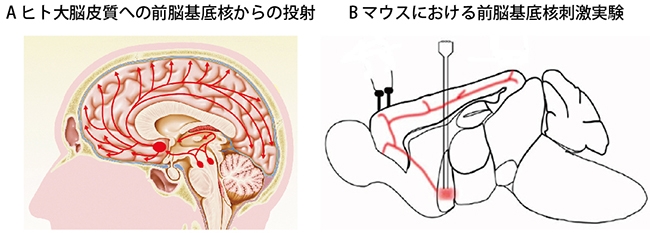
図1 アセチルコリン投射系とその刺激実験
- A.ヒト脳における前脳基底核から大脳皮質へのアセチルコリン投射系を示す。大脳基底部にある赤い楕円形部分がアセチルコリンを含む神経細胞の細胞体の存在する前脳基底核。
- B.マウスにおけるアセチルコリン投射線維(赤い線)と前脳基底核(赤くぼやけた四角)を示す。縦の2本線は前脳基底核刺激のため挿入した刺激電極を模式的に示したもの。左上の2本のスクリューは脳波記録のため留置した電極。(Alitto and Dan, Frontiers in Systems Neuroscience 6, Article 79,2013のFigure 1Aを改変)
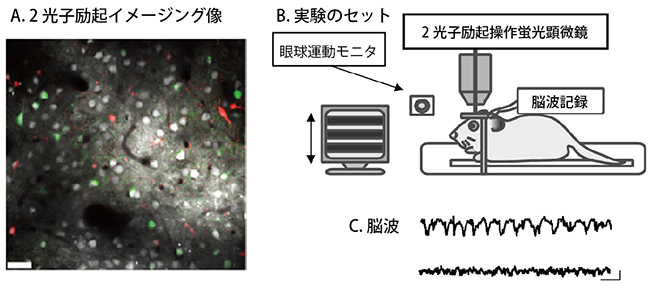
図2 ラット視覚野の神経細胞活動の2光子励起カルシウムイメージング法による記録
- A.大脳皮質表面より140μmの深さにおける各種細胞のイメージング像。白は興奮性神経細胞、緑は抑制性神経細胞、赤はグリア細胞を示す。左下のスケールバーは30μm。
- B.実験のセットの配置の模式図。眼前のディスプレイ画面上の縞が視覚刺激として種々の方向に動く。
- C.麻酔時の脳波(上)と覚醒時の脳波(下)の例。右下のスケールバーは5秒(横)と100μV(縦)。
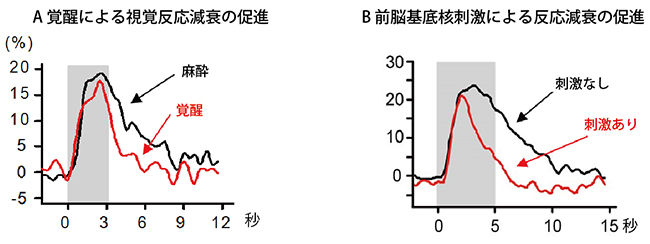
図3 覚醒による視覚反応と前脳基底核刺激による興奮性神経細胞の反応
- A.麻酔状態から覚醒状態になると、興奮性神経細胞は反応の減衰が速くなる。
- B.前脳基底核を電気刺激して視覚反応性の変化をみた結果。興奮性神経細胞の反応減衰の促進がみられる。
(グレーの部分は視覚刺激を与えた時間で縦軸は反応の強さを示す)
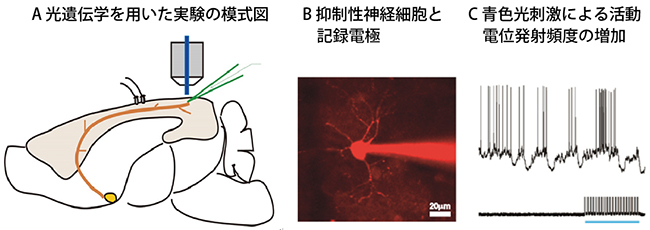
図4 光によるアセチルコリン投射系の刺激と大脳皮質抑制性神経細胞からの膜電位記録
- A.使用した遺伝子改変マウスは前脳基底核からの投射線維の末端でもチャンネルロドプシン2を発現しているため、青色光の照射によって活性化しアセチルコリンを放出する。大脳皮質神経細胞の反応はガラス管微小電極によって記録した。
- B.ガラス電極から注入された色素によって可視化された視覚野第1層の抑制性神経細胞。右下のスケールバーは20μm。
- C.青色光刺激を下の青横棒に示すように2秒間照射すると抑制性神経細胞の活動電位(上のトレースの縦線で示す)の発射頻度が増加する。
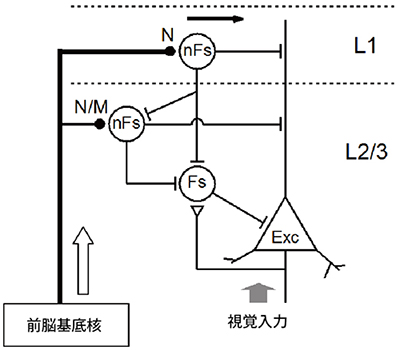
図5 覚醒による視覚反応の時間分解能の向上とその神経回路
覚醒によって前脳基底核が活動し、アセチルコリンが主に皮質第1層(L1)の抑制性神経細胞(nFs ;non-fast spiking cell)を活性化し、興奮性神経細胞(Exc)の視覚入力に対する反応を抑制する。ただ、この回路は時間遅れで働くため興奮性神経細胞の反応の減衰部分にのみ作用する。Nはニコチン受容体、Mはムスカリン受容体を示す。