ポイント
要旨
理化学研究所(理研)脳科学総合研究センター行動神経生理学研究チームの村山正宜チームリーダーらの国際共同研究グループ※は、皮膚感覚を知覚する脳の神経回路メカニズムを解明しました。
私たちが物に触れた時に得られる皮膚感覚の情報は、脊髄や視床を経由し大脳新皮質の第一体性感覚野(S1)[1]に到達した後、より高次な脳領域に伝わります。この低次領域から高次領域に向かう入力を「ボトムアップ入力」と呼びます。一方、高次から低次に向かう入力を「トップダウン入力」と呼びます。従来の仮説では、外界の情報に由来する外因性のボトムアップ入力と、注意や予測といった内因性のトップダウン入力とが脳のある領域で連合することで、皮膚感覚は知覚されると言われてきました。しかし、この仮説が正しければ、外因性のトップダウン入力だけでは皮膚感覚は知覚できず「注意して物を触らなければ何も感じない」ことになります。しかし、実際には特に注意せず、ぼーっとしている状態でも皮膚感覚の知覚は可能です。このように、皮膚感覚の知覚を形成する基本的な神経回路とそのメカニズムは全く分かっていませんでした。
国際共同研究グループは、マウスの肢を刺激した時に脳内で起こる神経活動を単一神経細胞レベルから回路レベルまで包括的に測定しました。また、マウスが皮膚感覚を識別する課題を行っている最中の行動を解析しました。その結果、内因性トップダウン入力と外因性ボトムアップ入力が同じタイミングで連合する神経活動は観察されませんでした。一方で、皮膚感覚の情報が外因性ボトムアップ入力としてS1から高次脳領域に送られた後、再びS1へ「外因性のトップダウン入力」として自動的にフィードバックされる反響回路[2]を発見しました。また、外因性トップダウン入力だけでも、従来提唱されてきた内因性トップダウン入力と外因性ボトムアップ入力の連合入力と同等な機能を担っていました。さらに、光遺伝学的手法[3]を用いてこの外因性トップダウン入力を抑制したところ、マウスは皮膚感覚を正常に知覚できなくなりました。
国際共同研究グループは、皮膚感覚の知覚における従来の神経回路モデルとは異なる新しいモデルを示しました。脳はこの2つの神経回路を状況により使い分けている可能性があります。神経科学者にとっての最重要課題の一つは、知覚などの「主観的な体験」を神経活動で説明することですが、本研究結果はその可能性を示しました。今後、詳細に外因性トップダウン入力のメカニズムを明らかすることで、老齢による五感の知覚能力の低下予防・改善の手がかりなどを得ることが期待できます。
本研究は、富士通株式会社及び株式会社富士通研究所の協力を得て実施しました。成果は、米国の科学雑誌『Neuron』(6月3日号)に掲載されるのに先立ち、オンライン版(5月21日付け:日本時間5月22日)に掲載されます。
※国際共同研究グループ
理化学研究所 脳科学総合研究センター
行動神経生理学研究チーム
チームリーダー 村山 正宜(むらやま まさのり)
客員研究員 真仁田 聡(まにた さとし)
研究員 鈴木 崇之(すずき たかゆき)
研究員 山田 一之(やまだ かずゆき)
訪問研究員 太田 桂輔(おおた けいすけ)(日本学術振興会 特別研究員)
テクニカルスタッフI 本間 千尋(ほんま ちひろ)
テクニカルスタッフI 松元 崇(まつもと たかし)
テクニカルスタッフI 小田川 摩耶(おだがわ まや)
テクニカルスタッフI 松原 智恵(まつばら ちえ)
記憶メカニズム研究チーム
チームリーダー 林 康紀(はやし やすのり)
客員研究員 佐藤 正晃(さとう まさあき)(JSTさきがけ研究員)
埼玉大学 理工学研究科/脳末梢科学研究センター
教授 中井 淳一(なかい じゅんいち)
准教授 大倉 正道(おおくら まさみち)
群馬大学 大学院医学系研究科 脳神経発達統御学講座 遺伝発達行動学分野
教授 柳川 右千夫(やながわ ゆちお)
名古屋大学 環境医学研究所 ストレス受容・応答研究部門 神経系分野Ⅱ
教授 山中 章弘(やまなか あきひろ)
特任助教 犬束 歩(いぬつか あゆむ)
フンボルト大学 神経科学研究センター
教授 Matthew Larkum(マシュー・ラーカム)
背景
私たちは五感を通して周りの世界を知覚しています。五感の中でも皮膚感覚(触覚)は、唯一、自分と相手に直接的かつ相互に影響する感覚です。目(視覚)、耳(聴覚)、鼻(嗅覚)、舌(味覚)からの情報は、体の局所的部位からの情報ですが、皮膚感覚は全身から得ることができます。物に触れた時に得られる皮膚感覚の情報は、脊髄や視床を経由し大脳新皮質の第一体性感覚野(S1)に到達することが、これまでの解剖学的・生理学的研究により知られています。S1に到達した情報は、より高次な脳領域に伝わります。このように低次領域から高次領域への入力を「ボトムアップ入力」と呼び、特に皮膚など感覚器からの外界入力を外因性ボトムアップ入力と呼びます。また反対に、高次領域から低次領域への入力を「トップダウン入力」と呼びます。
知覚に関する従来の仮説では、皮膚感覚を知覚するためには、皮膚からの外因性ボトムアップ入力とは別に、注意や予測といった脳内活動による内因性のトップダウン入力も関与すると考えられてきました。外因性ボトムアップ入力と内因性トップダウン入力が脳のある領域で連合することで皮膚感覚は知覚できると言われています。もしこの仮説が正しければ、私たちは「何かに注意しなければなにも感じない」ということにもなります。しかし、実際には私たちは、特に注意せずにぼーっとしている状態でも皮膚感覚を知覚することは可能であり、従来の仮説では私たちの実体験を説明できません。このように、これまで皮膚感覚の知覚を形成するための基本神経回路とそのメカニズムは分かっていませんでした。
そこで、国際共同研究グループはマウスを用いて皮膚感覚を知覚する脳内メカニズムを単一神経細胞レベルから回路レベルまで包括的に調べました。
研究手法と成果
国際共同研究グループは、マウスの皮膚感覚の知覚を司る神経回路を探るため、神経活動を広範囲に捉える膜電位イメージング[4]を大脳新皮質に対して行いました(図1A)。マウスの後肢を刺激すると、まず後肢に対応したS1の領域が活性化し、その後、第二運動野(M2)[5]が活性化しました(図1B)。次に神経活動を抑える薬をS1またはM2にそれぞれ投与し、その効果を観測しました。その結果、S1を抑制した場合はM2の活動が、逆にM2を抑制した場合はS1の遅い活動(遅発性神経活動)が抑制されました。これらの結果は、後肢からの情報はS1→M2→S1と流れることを示します(図1C)。これは、皮膚感覚が外因性ボトムアップ入力としてS1から高次脳領域であるM2に送られた後、再びS1へ「外因性のトップダウン入力」としてフィードバックされる反響回路が存在することを示します。
次に、反響回路における神経活動を詳細に調べるため、シリコン電極[6]を用いて活動電位応答を測定しました(図2A)。大脳新皮質は層構造をなしており、それぞれの層は異なる役割を担います。まず、S1における神経活動を皮質の全1-6層から記録したところ、全ての層において早発性神経活動と遅発性神経活動の2つのピークが記録されました(図2B)。一方、M2からの神経活動の記録では、S1のような2つのピークではなく1つのピークが観察されました。そのピークは、S1で観察された早発性神経活動と遅発性神経活動の間で見られました(図2C)。また、膜電位イメージングと同様にM2の活動を薬で抑制すると、S1の遅発性神経活動が有意に減少しました(図2D)。S1の早発性神経活動が外因性ボトムアップ入力、S1の遅発性神経活動がM2からの外因性トップダウン入力と考えられることから、膜電位イメージングで示された反響回路の存在を支持する結果と言えます。また、全6層のうち5層での遅発性神経活動が特に活発であることが分かりました(図2E)。これは、樹状突起が他細胞からの情報を受容し統合する役割を持つことから、全6層のうち最も長い樹状突起を持つ5層神経細胞では、より多くの情報を、より長い時間統合することが可能なためと考えられます。国際共同研究グループは、S1における5層での遅発性神経活動は、M2からの外因性トップダウン入力のみで引き起こされていることを解剖学的・生理学的に解明しました。5層での遅発性神経活動に対する外因性トップダウン入力単独が、従来提唱されている内因性トップダウン入力と外因性ボトムアップ入力の連合入力(図3A)と同等の機能を担うことが分かりました。この結果は、皮膚感覚の知覚における従来の神経回路モデルとは異なる新しい神経回路モデルを示したと言えます(図3B)。
さらに、国際共同研究グループは、光遺伝学的手法によりM2からS1への外因性トップダウン入力を特異的に抑制し、マウスの皮膚感覚の知覚における外因性トップダウン入力の役割を調べました。具体的には、マウスに対して皮膚感覚を手掛かりとする2種類の床面の識別する行動課題を行いました。1つ目の課題では、四角い箱の床面に紙やすり(ザラザラ)と、それを裏返した面(ツルツル)を半分ずつ敷き、その箱の中にマウスを置きました。マウスの脳には小型光刺激装置を設置し、外因性トップダウン入力を光刺激で抑制できるようにしました(図4A、B)。マウスはザラザラ、ツルツルの床面のどちらか一方を好む傾向があるため、光刺激をしないマウス群では、ザラザラ、またはツルツル床のどちらかに滞在時間が偏りました。しかし一方で、光刺激をしたマウス群では、その偏りが減少しました(図4C)。2つ目の課題では、Y字迷路の分岐点の手前の床面でザラザラ、またはツルツル床を提示し、ザラザラなら右方向、ツルツルなら左方向に進むよう訓練した後、光刺激の有無が正解率に与える影響を調べました(図4D)。光刺激をしないマウスは約80%の正解率を示しましたが、光刺激をしたマウスの正解率は約65%まで減少しました(図4E)。以上のことから、M2からS1への外因性トップダウン入力が、正常な皮膚感覚の知覚に必須であることが分かりました。
今後の期待
国際共同研究グループは、正常な皮膚感覚の知覚には、S1への外因性ボトムアップ入力だけでなく、その後のM2からS1への外因性トップダウン入力が必須であることを発見しました。また、今回発見した神経回路は、従来の回路とは異なり、注意をしなくても知覚できる回路として利用されている可能性があります。脳は2つの異なる神経回路を状態によって使い分けているかもしれません。神経科学者にとっての最重要課題の一つは、知覚などの「主観的な体験」を神経活動で説明することですが、本研究結果はその可能性を示しました。
また、脳卒中や事故などでの脳損傷が、体性感覚野ではなく高次領域に生じた場合でも半側空間無視などの失認[7]が起きますが、本研究結果はその一つのメカニズムとして、脳損傷によって高次領域がS1に外因性トップダウン入力を行えなくなっている可能性を示唆しています。今後、詳細にS1への外因性トップダウン入力のメカニズムを明らかすることで、失認の改善や老齢による五感の知覚能力の低下予防・改善の手がかりを得ることが期待できます。
原論文情報
- Satoshi Manita†, Takayuki Suzuki†, Chihiro Homma, Takashi Matsumoto, Maya Odagawa, Kazuyuki Yamada, Keisuke Ota, Chie Matsubara, Ayumu Inutsuka, Masaaki Sato, Masamichi Ohkura, Akihiro Yamanaka, Yuchio Yanagawa, Junichi Nakai, Yasunori Hayashi, Matthew E. Larkum & Masanori Murayama, "A Top-Down Cortical Circuit for Accurate Sensory Perception", Neuron, doi: 10.1016/j.neuron.2015.05.006
(†Equal contribution to this work)
発表者
理化学研究所
脳科学総合研究センター 行動神経生理学研究チーム
チームリーダー 村山 正宜(むらやま まさのり)
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.第一体性感覚野(S1)
大脳新皮質の頭頂葉に位置し、皮膚感覚や深部感覚をつかさどる低次脳領域。S1内の各所と体部位には対応関係がある。 - 2.反響回路
神経細胞集団Aと集団Bとが互いに出入力している回路。この回路では、集団Aの活動が集団Bの活動を増強し、その活動により再び集団Aが活性化する。実際の脳内では興奮性細胞だけでなく抑制性細胞活動も活性化されるので、いずれ神経活動は終息する。 - 3.光遺伝学的手法
神経回路機能を光と遺伝子操作を使って調べる手法。ミリ秒単位の時間精度をもった制御を特徴とする。本研究では光感受性抑制性イオンポンプ(ArchT)の遺伝子配列が組み込まれたアデノ随伴ウィルス(AAV)を脳内に注入することで、神経細胞にArchTを発現させた。その神経細胞に光を照射すると光感受性イオンポンプが活性化し、細胞内から細胞外にH+が排出され、結果的に神経細胞の活動は抑制される。 - 4.膜電位イメージング法
神経細胞の膜電位変化に応じて蛍光強度が変化する膜電位感受性色素で神経細胞を染色し、肢刺激で応答する神経活動を光学的に記録する。大脳新皮質表層(1-3層)の神経細胞の活動を広範囲に記録できる。 - 5.第二運動野(M2)
大脳新皮質の前頭野に位置し、動物行動の計画や実行、報酬や記憶に関連した活動を示す高次脳領域。 - 6.シリコン電極
本研究では16点の記録部位が1列に並んだ電極を用いた。この電極を大脳新皮質の層に対し垂直に刺入することで、神経活動を全層(1-6層)から同時に記録できる。 - 7.半側空間無視などの失認
半側空間無視は失認の一種で、体の左右どちらかからの感覚刺激(触覚、視覚、聴覚等)を知覚できなくなる症候のこと。大脳半球はその反対側の体部位の感覚を司っており、脳の障害部位により症候が異なる。
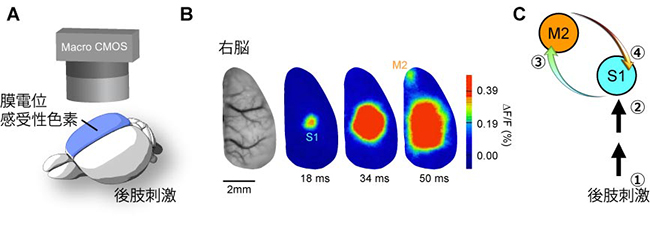
図1 第一体性感覚野(S1)と第二運動野(M2)間の反響回路の形成
- A: 大脳新皮質からの膜電位イメージング法の模式図。大脳新皮質に、神経応答に対応して蛍光強度変化を示す色素で新皮質を染色し、マクロ蛍光顕微鏡で神経応答を可視化した。
- B: マウスの後肢刺激で誘起される神経活動。まずS1で応答が起き、次にM2で生じた(注:その後M2はS1を再活性化する)。
- C: S1-M2反響回路。薬理実験(省略)で明らかになった情報の流れを番号で示す。①肢刺激、②視床を介したS1活動の誘起、③S1によるM2活動の誘起、④M2によるS1活動の再誘起
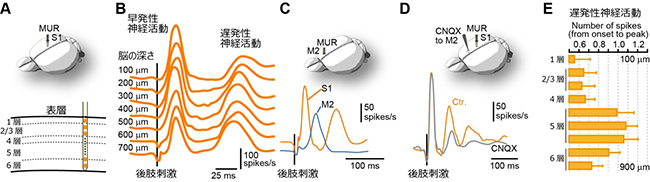
図2 S1とM2におけるマウス後肢刺激時の神経活動
- A: S1における全1-6層からの活動電位記録法(マルチユニット記録、MUR)の模式図。
- B: 後肢刺激で誘起された神経活動。早発性神経活動と遅発性神経活動が観察された。
- C: M2からの神経活動。M2の活動はS1における早発性神経活動と遅発性神経活動の間で生じた。
- D: M2の活動を薬で抑制するとS1における遅発性神経活動が減少した。
- E: 大脳新皮質各層における遅発性神経活動の大きさ。5層神経細胞が他層に比べより活性化していた。
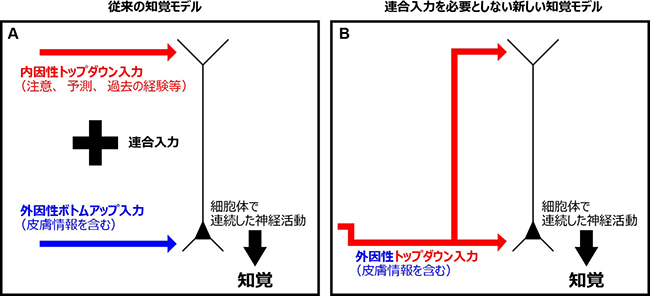
図3 従来の知覚モデルと新しいモデルとの違い
- A: 従来の知覚モデル。外因性ボトムアップ入力と内因性トップダウン入力との連合入力が神経細胞の樹状突起で統合され、細胞体では連続した活動電位の発生が生じる。その結果として皮膚感覚の知覚が生じるというモデル。
- B: 本研究結果により新たに提唱された、連合入力を必要としないモデル。S1→M2→S1の反響回路により皮膚情報を含む外因性トップダウン入力のみで、旧モデルと同様にS1神経細胞に連続した活動電位の発生を引き起こす。脳内ではAとB, 2つの異なる神経回路が脳の状態によって使い分けられている可能性がある。
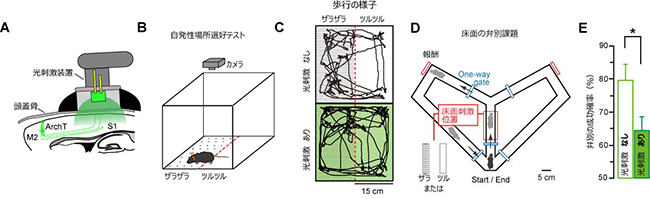
図4 M2からS1への外因性トップダウン入力が皮膚感覚の知覚に与える影響
- A: M2からS1への神経投射を特異的に抑制するための光刺激装置の模式図。M2には光感受性抑制性チャネルであるArchTを導入し、光刺激装置でM2からS1への外因性トップダウン入力を抑制した。
- B: 1つ目の課題の模式図。箱床に紙やすり(ザラザラ)と裏返した紙(ツルツル)を設置し、マウスがどちらにより長く滞在するかを記録した。
- C: 光刺激なし、ありのマウスにおける1つ目の課題の結果。光刺激なしの場合、マウスはどちらかの床に長く滞在する傾向を示した。一方、光刺激ありの場合、滞在時間の偏りが消失した。
- D: 2つ目の課題の模式図。Y字迷路の分岐点手前でザラザラまたはツルツル床をランダムに提示した。マウスは床面に合わせて右か左を選択し、正解であれば報酬を与えた。光刺激は床をマウスが通過する間に照射した。訓練を実施したのち、光刺激の有無が正解率に与える影響を調べた。
- E: 2つ目の課題の結果。光刺激なしの場合では、マウスは約80%の正解率を示すが、光刺激ありの場合、正解率が65%まで減少した。