要旨
理化学研究所(理研)環境資源科学研究センター植物ゲノム発現研究チームの関原明チームリーダー、松井章浩研究員らの共同研究チーム※は、アンチセンスRNA[1]による植物の新しい環境ストレス適応機構を発見しました。
植物の環境ストレスに対する適応機構の解明は、環境ストレスに対して強い作物を作出するために重要です。タンパク質をコードする遺伝子の環境ストレス下での転写制御について主要な部分は明らかになっていますが、環境ストレス適応における長鎖非翻訳型RNA[2]の機能および遺伝子の転写後制御機構についてはまだよく分かっていません。
今回、共同研究チームは、モデル植物のシロイヌナズナを用いて、環境ストレス下の植物体内で、1,000以上の遺伝子の長鎖非翻訳型アンチセンスRNAがRNA依存性RNAポリメラーゼ(RDR)[3]1/2/6により生成されることを明らかにしました。このアンチセンスRNAは、主にメッセンジャーRNA(mRNA)であるセンスRNAの分解産物を鋳型として作られ、センスRNAと2本鎖RNAを生成した後、small RNA[4]を生成せずにRNA分解を促進する新しい経路に含まれており、環境ストレス適応に関与することを見いだしました。
本研究を発展させることにより、植物の環境ストレスに対する遺伝子発現の制御機構の解明が進み、環境ストレス耐性作物の作出への利用が期待できます。
本成果は、米国の科学雑誌『Plant Physiology』オンライン版(7月14日付け)に掲載されました。
本研究は、日本学術振興会の科学研究費助成事業、科学技術振興機構(JST)戦略的創造研究推進事業チーム型研究(CREST)の支援を受けて実施されました。
※共同研究チーム
理化学研究所 環境資源科学研究センター 植物ゲノム発現研究チーム
チームリーダー 関 原明(せき もとあき)
研究員 松井 章浩(まつい あきひろ)
京都大学大学院医学研究科
特定助教 飯田 慶(いいだ けい)
基礎生物学研究所 生物機能解析センター
特任准教授 重信 秀治(しげのぶ しゅうじ)
背景
植物は動物と違い移動の自由がないため、乾燥などの環境ストレス変化に対して適応する独自の制御機構を備えています。植物の環境ストレスに対する応答機構の解明は、基礎研究および環境ストレス耐性作物の作出など応用の面からも重要です。
植物の環境ストレス応答におけるタンパク質をコードする遺伝子の転写制御機構について主要な部分は明らかになってきていますが、遺伝子の転写後の制御機構および長鎖非翻訳型RNAの役割についてはまだよく分かっていません。
関チームリーダーらは2008年、環境ストレス下で多数の遺伝子に非翻訳型アンチセンスRNAが存在することを報告しました注1)。これらのアンチセンスRNAの塩基配列はセンスメッセンジャーRNA(mRNA)の一部と一致していたことから、アンチセンスRNAはセンスRNAからRNA依存性RNA ポリメラーゼ(RDR)によって生成するのではないかと考えられました。しかし、アンチセンスRNAの生成メカニズムの詳細および環境ストレス適応における機能に関しては不明なままでした。
注1)2008年7月16日プレスリリース「約7,300種のシロイヌナズナ新規ノンコーディングRNAを同定」
研究手法と成果
モデル植物のシロイヌナズナのゲノムには、RDRをコードする六つの遺伝子が存在しています。共同研究チームはまず、RDR遺伝子の一重変異体および多重変異体などからアンチセンスRNAの蓄積量が減少する変異体を探索しました。その結果、rdr1/2/6三重変異体で1,000以上のタンパク質をコードする遺伝子領域において、ストレス誘導性のアンチセンスRNAの蓄積量が減少していることが分かりました。(図1)。さらに、分子機能を解析すると、ストレス誘導性のアンチセンスRNAはセンスRNAと2本鎖RNAを生成し、乾燥ストレス条件およびその後の再給水による回復過程において、センスmRNAの分解速度が野生株と比べて減少していました(図2)。また、アンチセンスRNAの蓄積量は、キャップ構造[5]の除去に関与するDCP5遺伝子の変異体で減少し、5’末端からのRNA分解に関与するXRN4遺伝子の変異体で増加していました。
これらの結果から、ストレス誘導性のアンチセンスRNAは、センスRNAの分解産物を鋳型としてRDR1/2/6により生成されると考えられます。rdr1/2/6三重変異体は、乾燥ストレス処理後の回復過程で根の成長が野生株と比べて抑えられており(図3)、アンチセンスRNAが環境ストレス適応において生理的に重要な役割を果たしていることが示されました。
一方、以前の研究でRDRは特殊な条件でタンパク質をコードする遺伝子領域からmRNAの分解産物を使ってsmall RNAを生成することが報告されています。しかし予想に反して、共同研究チームが発見したストレス誘導性のアンチセンスRNAが生成する遺伝子領域では、small RNAの生成がほとんど観察されませんでした(図4)。さらに、既知のsmall RNA生成経路に関与する変異体dcl2/3/4において、ストレス誘導性アンチセンスRNAの蓄積量、センスRNAの分解速度、根の伸長などは野生株との間に違いは観察されませんでした。
以上の結果から、RDR1/2/6により生成されるストレス誘導性の非翻訳型アンチセンスRNAは、既知のsmall RNA生成経路とは異なる新しいセンスRNA分解機構に含まれ、環境ストレス適応に関与することが分かりました(図5)。
今後の期待
本研究からこれまで知られていたsmall RNAの生成経路とは異なる、非翻訳型アンチセンスRNAによるセンスRNAの新しい分解経路が環境ストレス適応において機能することが分かりました。このことからRNA分解は予想以上に複雑な機構によって制御されていることがみえてきました。また、mRNAの分解制御[6]が環境ストレス下で一度停止した成長が再開する過程で重要な役割を持つことも新たに分かりました。
本結果によって植物の環境ストレスに対する新たな遺伝子発現の制御機構の解明が進み、環境ストレス耐性作物の作出への利用が期待できます。
原論文情報
- Akihiro Matsui, Kei Iida, Maho Tanaka, Katsushi Yamaguchi, Kayoko Mizuhashi, Jong-Myong Kim, Satoshi Takahashi, Norio Kobayashi, Shuji Shigenobu, Kazuo Shinozaki, and Motoaki Seki, "Novel stress-inducible antisense RNAs of protein-coding loci are synthesized by Arabidopsis RDRs", Plant Physiology, doi: 10.1104/pp.17.00787
発表者
理化学研究所
環境資源科学研究センター 植物ゲノム発現研究チーム
チームリーダー 関 原明(せき もとあき)
研究員 松井 章浩(まつい あきひろ)
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
お問い合わせフォーム
産業利用に関するお問い合わせ
理化学研究所 産業連携本部 連携推進部お問い合わせフォーム
補足説明
- 1.アンチセンスRNA
タンパク質をコードするメッセンジャーRNA(mRNA)のように意味を持つ配列として転写されたRNAをセンスRNAと呼ぶのに対し、その相補的な配列を持つRNAをアンチセンスRNAと呼ぶ。 - 2.非翻訳型RNA
タンパク質をコードしないDNA配列から転写され、タンパク質に翻訳されないRNA。ノンコーディングRNA(ncRNA)ともいう。200塩基以上の非翻訳型RNAを特に長鎖非翻訳型RNAと呼ぶことがある。 - 3.RNA依存性RNAポリメラーゼ(RDR)
RNAを鋳型としてRNAを合成する酵素。 - 4.small RNA
タンパク質に翻訳されない20~30塩基程度の短いRNA。 - 5.キャップ構造
mRNAの5’末端には7-メチルグアノシンが付加されており、これをキャップ構造という。キャップ構造はタンパク質合成に必須の構造である。mRNAはキャップに結合する特殊なタンパク質を介してリボソームと結合する。 - 6.mRNAの分解制御
mRNAの蓄積量は、遺伝子の転写速度とmRNAの分解速度のバランスにより制御されている。近年、mRNA分解を介した遺伝子発現制御の重要性が明らかになっている。
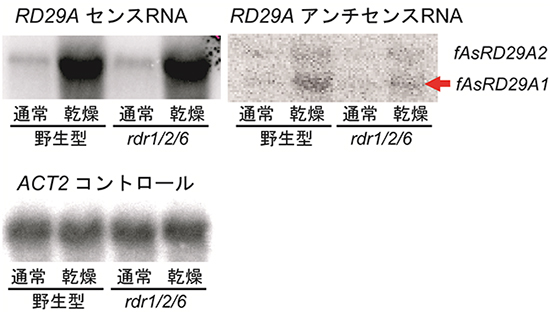
図1 シロイヌナズナのrdr1/2/6三重変異体におけるアンチセンスRNA蓄積量の減少
シロイヌナズナrdr1/2/6変異体におけるRD29A遺伝子領域のアンチセンスRNAの発現をノーザンブロット法により測定した。RDR1/2/6依存性アンチセンスRNAを赤矢印で示している。rdr1/2/6三重変異体では、乾燥ストレス条件でアンチセンスRNA発現量が減少している。
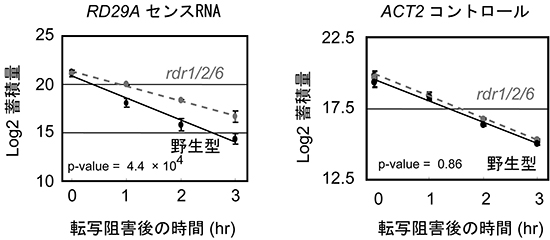
図2 rdr1/2/6三重変異体における転写阻害後のセンスRNAの分解速度
転写阻害剤を使って新たなRNAの転写を阻害し、RD29AセンスRNAの分解速度を測定した。rdr1/2/6変異体では、RD29AセンスRNAの分解速度が野生株と比べて減少した。

図3 rdr1/2/6三重変異体における乾燥ストレス処理後の回復過程での根の成長
水が飽和している状態(左)と水を切って3回乾燥ストレスを与えたシロイヌナズナ(右)。3回乾燥ストレスを与えたrdr1/2/6変異体は、野生株と比較して根の成長が抑えられていた。
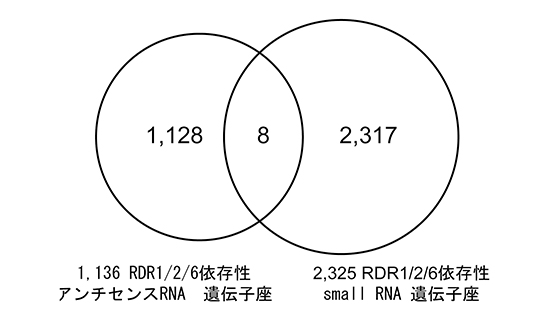
図4 アンチセンスRNA遺伝子座とsmall RNA遺伝子座の非重複
RDR1/2/6依存性アンチセンスRNAを持つ遺伝子座とRDR1/2/6依存性small RNAを持つ遺伝子座を比較した。ほとんど重複する遺伝子がないことから、RDR1/2/6依存性のアンチセンスRNAは既知のsmall RNA生成経路と関連していないといえる。
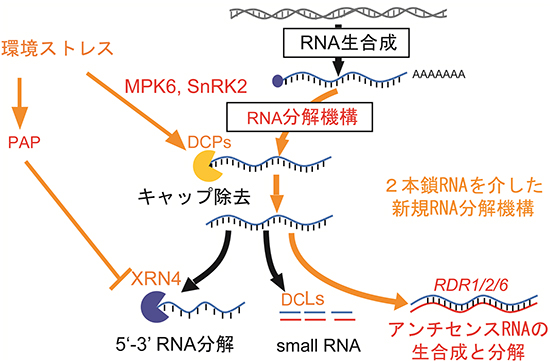
図5 環境ストレス下でのアンチセンスRNAによるセンスRNA分解のモデル図
アンチセンスRNAの蓄積量は、キャップ構造の除去に関与するDCP5遺伝子の変異体で減少し、5’末端からのRNA分解に関与するXRN4遺伝子の変異体で増加していることから、センスRNAの分解産物を鋳型としてRDR1/2/6により生成していると考えられた。また、small RNAも生成されていないことから、これまで知られているsmall RNAの生成経路とは異なり、アンチセンスRNAはセンスRNAと2本鎖RNAを生成してセンスRNAを分解する新しい経路を発見した。