2010年5月13日
独立行政法人 理化学研究所
再生阻害因子が神経細胞の突起をはねのける仕組みを発見
-神経回路の構築や損傷後の再生に重要な役割を果たす新メカニズム-
ポイント
- 神経突起の表面の膜が、細胞内へ取り込まれる様子を初観察
- 再生阻害因子が、形質膜の細胞内への取り込みを促進し、神経突起をはねのける
- 損傷した神経回路の修復技術への応用に期待
要旨
独立行政法人理化学研究所(野依良治理事長)は、神経回路の再生を妨げる分子の作用機序を解析し、これらの再生阻害因子が神経突起※1の形質膜※2の取り込みを促進し、神経突起をはねのけることを世界で初めて発見しました。理研脳科学総合研究センター(利根川進センター長)神経成長機構研究チームの上口裕之チームリーダー、戸島拓郎研究員、糸総るり香テクニカルスタッフによる研究成果です。
脳や脊髄の働きの中心を担う神経回路は、多数の神経細胞から伸びた突起が精巧に絡み合って構築されています。この神経回路の構築には、成長円錐※3と呼ばれる神経突起の先端部が、その周囲環境に存在する分子情報を感受して、神経突起を正しい方向へ導くことが必要です。神経回路が損傷すると、神経突起の切断端そのものが新たに成長円錐となり、その神経突起が伸長して神経回路を再構築しようとします。しかし、損傷組織由来の再生阻害因子が成長円錐を反発するため、神経突起は損傷部位を通過することができません。このため、損傷した神経回路の修復技術を開発するためには、再生阻害因子が成長円錐を反発する仕組みを理解することが必須となっています。
研究チームは、神経細胞の培養液中に再生阻害因子の濃度勾配を作り、この培養液の環境を移動する成長円錐の形質膜の取り込み(クラスリン依存性エンドサイトーシス※4)を解析しました。すると、高濃度の再生阻害因子に面した成長円錐の片側でクラスリン依存性エンドサイトーシスが亢進し、その結果、成長円錐は反対側に旋回して再生阻害因子を避ける方向へ移動していきました。クラスリン依存性エンドサイトーシスを抑制すると、成長円錐は、逆に再生阻害因子に誘引される方向に旋回しました。このことから、再生阻害因子は、成長円錐でのクラスリン依存性エンドサイトーシスを非対称化することで、神経突起を反発することが判明しました。
本研究は、成長円錐での非対称性エンドサイトーシスが、神経突起の伸長方向を決定するという基本原理を明らかにし、神経突起の再生を可能とする新たな分子メカニズムを発見した意義のある成果となりました。
本研究成果は、米国の科学雑誌『Neuron(ニューロン)』(5月13日号)に掲載されます。成果の一部は科学研究費補助金を得て実施しました。
背景
私たちは、神経回路を使って感覚、運動、記憶、学習などさまざまな活動を営んでいます。この神経回路は、多数の神経細胞から伸びた突起が精巧に絡み合った連絡網ですが、どのような仕組みで連絡網を形成しているのか、どのような方法で傷ついた連絡網を修復しているのかは、まだ十分に解明されていません。伸長途上の神経突起の先端部は、成長円錐と呼ばれるアメーバ状に広がった運動性に富む構造をしており(図1A)、周囲の細胞外環境に存在するガイダンス分子※5を感受して、神経突起を正しい方向へ伸ばします。多くのガイダンス分子は、成長円錐から離れた場所にある細胞から産生・放出されて、成長円錐近傍の細胞外に濃度の勾配を作ります。ガイダンス分子には成長円錐を誘引する因子(誘引性ガイダンス分子)と反発する因子(反発性ガイダンス分子)があります。ガイダンス分子は、成長円錐の表面に存在する受容体に結合し、細胞質カルシウムイオン(Ca2+)シグナル※6を生成して、最終的に誘引性ガイダンス分子は神経突起を誘引し、反発性ガイダンス分子は神経突起を反発します。成長円錐の片側に、より多く結合したガイダンス分子は、同じ側の細胞質でCa2+濃度を上昇させて成長円錐の移動方向を転換します。すなわち、成長円錐の片側(ガイダンス分子に遭遇した側)で発生したCa2+シグナルが、成長円錐を誘引または反発する方向に旋回させることができます。
研究チームはこれまでに、成長円錐を誘引するCa2+シグナルと反発するCa2+シグナルの性質の違いを明らかにしています。これらのCa2+シグナルを人工的に生成する手法を確立し、誘引性Ca2+シグナルに応答して成長円錐が旋回するメカニズムも解明しました(2006年12月11日プレスリリース)。しかし、反発性Ca2+シグナルが成長円錐の旋回を駆動する仕組み、すなわち反発性ガイダンス分子が成長円錐を反発する細胞内メカニズムは未解決の課題でした。
神経回路が断裂すると、神経突起の切断端が新たに成長円錐となり、その成長円錐は再び移動して神経突起を再生しようとします。しかし、損傷した中枢神経組織が産生する再生阻害因子が、成長円錐をはねのけるため、神経突起は損傷部位を通過することができず、神経回路は修復されません。再生阻害因子の多くは、反発性ガイダンス分子です。また、再生阻害と反発性ガイダンスの間には、共通のメカニズムが多いことも知られています。このため、反発性ガイダンスの仕組み、すなわち再生阻害因子が成長円錐をはねのける仕組みを明らかにすることは、神経回路の修復技術を開発するために必要不可欠な知見です。そこで研究チームは、代表的な再生阻害因子であるセマフォリン3A(Sema3A)が成長円錐を反発するメカニズム、特に反発性Ca2+シグナルが成長円錐の旋回を駆動する仕組みを解明することに挑みました。
研究手法と成果
研究チームは、成長円錐での形質膜の取り込みを可視化するため、クラスリン依存性エンドサイトーシスにかかわるタンパク質(クラスリンとダイナミン)の挙動を全反射蛍光顕微鏡※7で観察しました(図1B)。その結果、成長円錐の形質膜の一部がクラスリンの働きで細胞内に向かって陥入し、この陥入した形質膜をダイナミンが引きちぎり、引きちぎられた膜が細胞内へ取り込まれる過程(クラスリン依存性エンドサイトーシス)を捉えることに成功しました(図2)。
次に、誘引性あるいは反発性Ca2+シグナルが、クラスリン依存性エンドサイトーシスに及ぼす影響を解析するため、上下2本の対物レンズで培養細胞を挟み込む特殊な顕微鏡システムを構築しました。この顕微鏡システムの上側のレンズから成長円錐に紫外線を照射して、反発性および誘引性のCa2+シグナルを人工的に作り出し、下側のレンズ(全反射蛍光顕微鏡の一部)でクラスリン依存性エンドサイトーシスを観察しました。成長円錐の片側に反発性Ca2+シグナルを生成すると、クラスリン依存性エンドサイトーシスの頻度が約1.5倍に亢進しましたが、Ca2+シグナルが生成している反対側ではエンドサイトーシスは変化しませんでした(図3)。一方、成長円錐に誘引性Ca2+シグナルを生成しても、クラスリン依存性エンドサイトーシスは変化しませんでした。この実験で、成長円錐の反発性Ca2+シグナルはクラスリン依存性エンドサイトーシスを非対称化することが分かりました。
このような非対称性エンドサイトーシスが、成長円錐の反発に必要か否かを検証するため、各種薬剤や変異型タンパク質でクラスリン依存性エンドサイトーシスを抑制した成長円錐の旋回を解析しました。エンドサイトーシスを抑制した成長円錐は、誘引性Ca2+シグナルに反応して旋回することはできますが、反発性Ca2+シグナルに反応して旋回することはできませんでした(図4)。これにより、反発性Ca2+シグナルによる成長円錐の旋回は、非対称的なクラスリン依存性エンドサイトーシスを必要とすることが判明しました。
Sema3Aなどの再生阻害因子は、成長円錐に反発性Ca2+シグナルを生成して、神経突起を反発します。そこで研究チームは、再生阻害因子が成長円錐を反発する過程で、Ca2+シグナルの下流で非対称性エンドサイトーシスが関与するか否かを検証しました(図5A)。成長円錐の近傍の培養液中にSema3Aの濃度勾配を作ると、高濃度のSema3Aに面した片側でクラスリン依存性エンドサイトーシスが亢進し(図5B)、成長円錐はSema3Aを避けて濃度の低い方向へ旋回しました(図5C)。薬剤処理でクラスリン依存性エンドサイトーシスを抑制した成長円錐は、Sema3Aに反発されずに、逆にSema3Aに誘引される方向に伸長しました(図5D)。これにより、再生阻害因子はクラスリン依存性エンドサイトーシスを非対称化して成長円錐を反発することと、エンドサイトーシスを抑制した神経突起は再生阻害因子に向かって伸長できることが明らかになりました。
今後の期待
研究チームは、2006年に誘引性ガイダンスのメカニズムを発表しました。誘引性Ca2+シグナルは、成長円錐でのエキソサイトーシス※8を非対称化し、Ca2+シグナル発生側の形質膜に新たな膜成分を供給することで、成長円錐の旋回を駆動します。今回の研究成果は、エキソサイトーシスとエンドサイトーシスという相反する膜動態のバランスが神経突起の伸長方向を決定するという基本原理を明らかにし(図6)、神経回路構築の研究に影響を及ぼす重要な概念を示しました。特に、エンドサイトーシスを抑制した神経突起が再生阻害されなくなるという発見は、神経再生分野への貢献が期待できます。今後は、再生阻害因子/反発性ガイダンス分子が成長円錐の膜動態を制御するメカニズムを詳細に研究し、生体内で損傷した神経回路の修復技術への応用を目指します。
発表者
理化学研究所
脳科学総合研究センター 疾患メカニズムコア 神経成長機構研究チーム
Tel: 048-467-6137 / Fax: 048-467-9795
お問い合わせ先
脳科学研究推進部Tel: 048-467-9757 / Fax: 048-462-4914
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.神経突起
神経細胞から伸びる細長い突起で、遠隔部位に存在するほかの神経細胞や筋肉細胞などとシナプスを形成して、神経回路を構築する。 - 2.形質膜
細胞の表面を構成する膜構造。 - 3.成長円錐
発生再生過程の脳神経組織で、伸長している神経突起の先端部に形成される、アメーバ状の形をした構造体。細胞外に存在する標識(ガイダンス分子など)を感知し、正しい道のりを移動することにより、神経突起を目的の方向へ誘導する。 - 4.クラスリン依存性エンドサイトーシス
細胞の内部(細胞質)に存在するクラスリンやダイナミンなどの働きにより、形質膜の一部が細胞内へ取り込まれること。クラスリンに裏打ちされた形質膜が細胞質側へ陥入し、この陥入部分がダイナミンの働きにより形質膜から離断し、膜成分とそれに結合した細胞外物質が細胞内へ取り込まれる。 - 5.ガイダンス分子
神経突起が伸長する道筋付近に提示され、成長円錐の移動方向を制御する分子群。 - 6.細胞質カルシウムイオン(Ca2+)シグナル
細胞質のカルシウムイオン(Ca2+)が関与する情報伝達のこと。Ca2+濃度の上昇で、さまざまな分子の働きを制御し、細胞機能に影響を及ぼす。 - 7.全反射蛍光顕微鏡
培養細胞が付着したカバーガラスに斜めにレーザー光を照射して、カバーガラス表面から培養細胞に向かって約100ナノメートルの範囲を選択的に観察するための顕微鏡。細胞のカバーガラスへの付着面(形質膜とその近傍)での分子の挙動を捉えるのに適した観察方法。 - 8.エキソサイトーシス
細胞内にある膜成分を形質膜と融合・一体化すること。
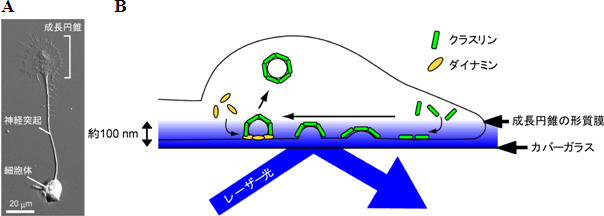
図1 全反射蛍光顕微鏡によるクラスリン依存性エンドサイトーシスの観察方法
(A)神経突起と成長円錐の形態。
(B)カバーガラス上の成長円錐を横方向から見た模式図。成長円錐の形質膜の一部が、クラスリンに裏打ちされて細胞内へ陥入し、ダイナミンによって引きちぎられて取り込まれる様子を示す。全反射蛍光顕微鏡を用いて、カバーガラス上面から約100ナノメートルの範囲でのクラスリンとダイナミンの挙動を解析した。
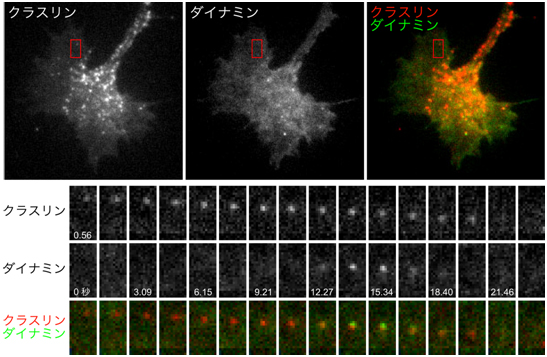
図2 全反射蛍光顕微鏡によるクラスリン依存性エンドサイトーシスの観察像
上段の3枚の画像は、成長円錐の形質膜近傍のクラスリンとダイナミンおよび両者の重ね合わせ像を示す。赤線で囲った長方形の領域を拡大し、経時的な変化を下段に示した(数字は経過した秒数を表す)。クラスリンが形質膜を裏打ちし(0秒付近で出現したクラスリンの斑点)、同部位にダイナミンが一時的に集積し(15秒付近)、クラスリンに覆われた膜は形質膜から離断して細胞内へ移動するため、全反射蛍光顕微鏡では見えなくなる(21秒付近)。
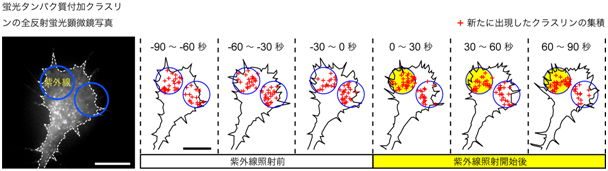
図3 反発性Ca2+シグナルによるクラスリン依存性エンドサイトーシスの非対称化
成長円錐の片側(成長円錐の左側の青丸の領域)に紫外線を照射して、反発性Ca2+シグナルを生成した。紫外線は、3秒ごとに100ミリ秒間照射した。成長円錐に発現する蛍光タンパク質付加クラスリンの挙動を全反射蛍光顕微鏡で撮影し、紫外線照射の開始前後の経時変化を解析した。成長円錐両側の青丸の領域内に出現したクラスリンの斑点(クラスリン依存性エンドサイトーシスの開始に相当)を30秒毎に数え、斑点の出現部位を+印で図示した。反発性Ca2+シグナルが発生した領域(黄色)でのクラスリン依存性エンドサイトーシスの頻度は、反対側の青丸の領域での頻度と比較して約1.5倍であった。スケールバーは10μmに相当。
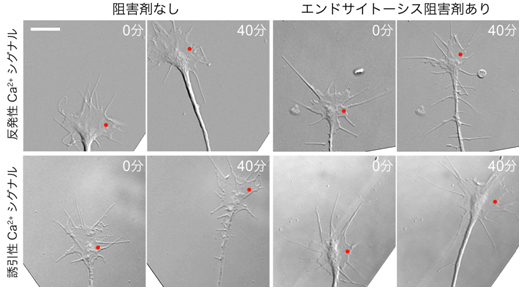
図4 クラスリン依存性エンドサイトーシスは成長円錐の反発に必要
成長円錐の片側(赤点)に紫外線を照射して、反発性Ca2+シグナル(上段)あるいは誘引性Ca2+シグナル(下段)を生成し、成長円錐の旋回を誘起した。紫外線照射から40分後、反発性Ca2+シグナルを生成した成長円錐は反発方向に(左上写真2枚)、誘引性Ca2+シグナルを生成した成長因子は誘引方向に(左下写真2枚)旋回した。紫外線照射開始前にクラスリン依存性エンドサイトーシスを阻害する薬剤を投与しておくと、成長円錐は反発性Ca2+シグナルに応答せずに直進したが(右上写真2枚)、誘引性Ca2+シグナルには応答して旋回した(右下写真2枚)。スケールバーは10μmに相当。
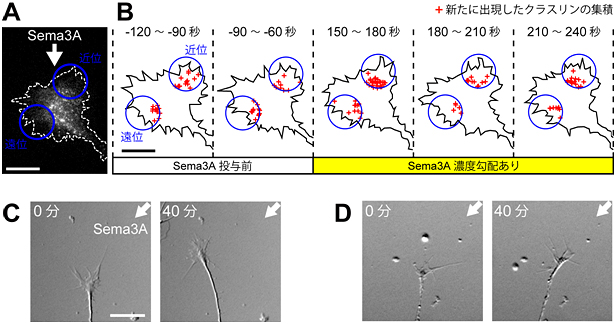
図5 Sema3Aは成長円錐でのエンドサイトーシスを非対称化して神経突起を反発する
(A)蛍光タンパク質付加クラスリンを発現する成長円錐を全反射蛍光顕微鏡で撮影した。成長円錐の斜め前方(矢印の方向)からSema3Aを投与して、培養液中にSema3Aの濃度勾配を作製した。スケールバーは10μmに相当。
(B)Sema3Aの濃度勾配を作製する前後でのクラスリン依存性エンドサイトーシスの経時変化を解析した。成長円錐の両側に青丸の領域を設定し、30秒間毎のクラスリン斑点の出現部位を+印で図示した。高濃度Sema3Aに面する近位側でのクラスリン依存性エンドサイトーシスの頻度は、遠位側での頻度と比較して約2倍であった。スケールバーは10μmに相当。
(C)Sema3A投与開始時と40分経過後を観察した。矢印の方向からSema3Aを投与すると、成長円錐を反発し、神経突起はSema3A濃度勾配を避ける方向に伸長した。スケールバーは20μmに相当。
(D)あらかじめ培養液全体にクラスリン依存性エンドサイトーシスの阻害剤を添加しておき、その後に矢印の方向からSema3Aの投与を開始した。Sema3A投与開始時と40分経過後の写真を示す。エンドサイトーシスを抑制した神経突起はSema3A濃度の高い方向に伸長した。
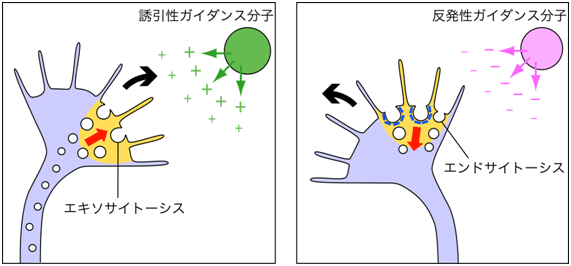
図6 成長円錐での非対称性膜動態は誘引性/反発性ガイダンスを駆動する
誘引性/反発性ガイダンス分子の濃度勾配に遭遇した成長円錐は、遭遇側にCa2+シグナル(橙色)を発生する。誘引性ガイダンス分子と遭遇して誘引性Ca2+シグナルが発生すると(左図)、細胞内の膜成分が成長円錐の先端部へ運ばれ、エキソサイトーシスを起こして形質膜に供給される。一方、反発性ガイダンス分子と遭遇して反発性Ca2+シグナルが発生すると(右図)、形質膜の一部がクラスリン依存性エンドサイトーシスによって除去される。このように、成長円錐の片側での膜の供給と除去が、旋回を駆動する。