2013年10月1日
独立行政法人理化学研究所
独立行政法人科学技術振興機構
細胞の運命を左右する新しい分子メカニズムの一端を解明
-ポリコム複合体間で起こる重合が遺伝子発現のオン・オフを調節-
ポイント
- 遺伝子制御に関わるポリコム複合体のライブイメージングに成功
- ポリコム複合体が重合し、ポリコム構造体を形成
- ポリコム複合体重合の制御メカニズム解明が再生医療やがん治療に貢献
要旨
理化学研究所(理研、野依良治理事長)は、細胞の運命を左右する新しい分子メカニズムの一端を解明しました。これは理研統合生命医科学研究センター(小安重夫センター長代行)免疫器官形成研究グループの古関明彦グループディレクター、磯野協一上級研究員(JST戦略的創造研究推進事業さきがけ研究者兼務)らの研究チームの成果です。
細胞の運命は、幹細胞など多能性を維持した未分化細胞の時期に決定されます。その時期に、運命決定に関わるさまざまな遺伝子のオン・オフが切り換わります。この遺伝子発現のオン・オフの切り換えを管理しているのが、ポリコムタンパク質群[1]です。ポリコムタンパク質群は、その標的となる遺伝子の特定の部分で巨大な複合体(ポリコム複合体)を形成し、標的遺伝子を不活性化することが知られていましたが、その分子メカニズムは解明されていませんでした。
研究チームは、遺伝子改変技術と細胞イメージング手法を使い、ポリコム複合体が不活性状態の標的遺伝子上で重合し、「ポリコム構造体」を形成することを発見しました。ポリコム構造体の役割を理解するために、その構成成分の1つである「Phc2タンパク質」が持つ自己重合活性[2]に注目しました。自己重合不全を引き起こすPhc2遺伝子点変異[3]マウスを作製し解析したところ、点変異細胞では、ポリコム構造体の消失、クロマチン[4]構造の緩み、さらにポリコム複合体の制御下にある遺伝子群の発現上昇が起こりました。これらの異常によって体節の運命が変化し、本来は頸椎となるべき脊椎が胸椎の特性を持つようになりました。これらの結果から、ポリコム複合体は遺伝子抑制機能を発揮するためにPhc2タンパク質によって重合し、ポリコム構造体を形成することが明らかになりました。一方、遺伝子発現の切り換えの観点から推察すると、遺伝子活性化のためにポリコム複合体の重合を解く仕組みも備わっていると考えられます。
今後、細胞内外の刺激やストレスに応答したポリコム複合体重合を制御するメカニズムの解明は、再生医療やがん治療に役立つと期待できます。本研究は、科学技術振興機構(JST)戦略的創造研究推進事業 個人型研究(さきがけ)の一環として行われました。成果は、米国の科学雑誌『Developmental Cell』(9月30日号)に掲載されます。
背景
未分化状態の幹細胞から特定の細胞へと分化する細胞の運命決定には、どの遺伝子をどこで発現(オン)させるか、あるいは抑制(オフ)するかといった遺伝子発現の切り換えが深く関わっています。幹細胞が分化するための多能性を維持するには、分化に関連する遺伝子が活性化できる性質を保ちながらも実際には不活性な状態に留まっている必要があります。同時に幹細胞は、分化などで減少する分を補うために自己複製を繰り返しています。
このような幹細胞の特性を維持するために、重要な役割を果たしているのがポリコムタンパク質群です。ポリコムタンパク質群は、細胞の分化や増殖抑制に関わる遺伝子領域に複合体(ポリコム複合体)を形成して、その遺伝子を不活性化することが報告されています。従って、ポリコム複合体は、幹細胞を起点する細胞の運命決定の主要因子といえます。しかし、ポリコム複合体がどのように遺伝子を抑制し、また、どのようにその抑制を解除するのか、その分子メカニズムは、ほとんど解明されていませんでした。
研究手法と成果
研究チームは、ポリコム複合体による遺伝子制御メカニズムを理解するために、マウス遺伝子改変技術と細胞イメージング技法を併用しました。まず、ポリコム複合体の核内局在を知るために、ポリコムタンパク質を蛍光で追跡できる遺伝子改変マウスを作製しました。改変マウス胚由来の細胞を顕微鏡観察したところ、不活性化状態の遺伝子領域(Hox遺伝子[5]など)でポリコム複合体が重合して連なり、斑点状の「ポリコム構造体」を形成することが分かりました(図1)。
ポリコム構造体の役割を適切に理解するためには、構造体形成を阻害する必要がありました。研究チームは、ポリコム複合体の構成成分の1つ「Phc2タンパク質」が持つ自己重合活性がカギになっていると考えました。そこで、重合形成不全を引き起こす遺伝子変異マウス(Phc2遺伝子点変異マウス)を作製し、細胞を観察しました。その結果、予想した通りにポリコム構造体は消失しました(図2)。また、ポリコム構造体が形成されてない状態では、遺伝子発現のオン・オフに深く関わっているクロマチン構造が緩むとともに、ポリコム複合体の制御下にある遺伝子群(Hox遺伝子、がん抑制遺伝子Cdkn2a[6]など)の発現が上昇しました(図3、4)。Hox遺伝子の発現上昇は、脊椎の形成パターンに影響を及ぼすことが知られています。また、Cdkn2a遺伝子の発現上昇は、早期細胞老化[7]を引き起こします。実際にポリコム構造体不全がこのような現象を誘導するかを調べました。その結果、Phc2遺伝子点変異マウスでは、本来は頸椎になるべき脊椎が胸椎に、胸椎になるはずの脊椎が腰椎になるという体節の運命変換が起こっていました(図5)。加えて、Phc2点変異細胞は早期に老化することも確認しました。以上の結果は、Phc2タンパク質の重合活性によって起こるポリコム複合体の重合、言い換えればポリコム構造体の形成が細胞の運命決定に関わる遺伝子の抑制に必要であることを示しています(図6)。
今後の期待
今回の研究成果は、ポリコム複合体の重合の促進と解除が遺伝子発現のオン・オフの切り換えに直結している可能性を示しています(図6)。今後、この制御メカニズムを解明できれば、多能性を持つ胚性幹細胞(ES細胞)や人工多能性幹細胞(iPS細胞)の運命コントロールが可能となり、再生医療の進展に役立つと考えられます。また、近年、ポリコム複合体の過剰機能ががん化の主要原因の1つとなっていることが明らかにされています。ポリコム複合体の重合を制御するメカニズムの解明は、がん化の抑止、あるいはがん細胞を死滅させる抗がん剤開発に役立つ可能性があります。
ポリコム構造体はポリコム複合体の機能を反映したものです。今後、ポリコム構造体の形態変化を詳しく調べることで、再生医療やがん治療に応用できるポリコム複合体の重合による遺伝子発現のオン・オフの切り換えの核心に迫れると期待しています。
なお、本研究はJST 戦略的創造研究推進事業 個人型研究(さきがけ)「エピジェネティクスの制御と生命機能」研究領域(研究総括:向井常博 西九州大学学長)における研究課題「細胞運命に関わるポリコーム群制御の切り換え機構」(研究者:磯野協一)の一環として行われました。
原論文情報
- Kyoichi Isono, Takaho A. Endo, Manching Ku, Daisuke Yamada, Rie Suzuki, Jafar Sharif, Tomoyuki Ishikura, Tetsuro Toyoda, Bradley E. Bernstein, and Haruhiko Koseki. "SAM domain polymerization links subnuclear clustering of PRC1 to gene silencing" Developmental Cell, 2013, http://dx.doi.org/10.1016/j.devcel.2013.08.016
発表者
理化学研究所
統合生命医科学研究センター 免疫器官形成研究グループ
上級研究員 磯野 協一(いその きょういち)
お問い合わせ先
統合生命医科学研究推進室
金子 明義(かねこ あきよし)
Tel: 045-503-9121 / Fax: 045-503-9113
報道担当
独立行政法人理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
独立行政法人科学技術振興機構 広報課
Tel: 03-5214-8404 / Fax: 03-5214-8432
JSTの事業に関すること
独立行政法人科学技術振興機構 戦略研究推進部
Tel: 03-3512-3525 / Fax: 03-3222-2063
補足説明
- 1.ポリコムタンパク質群
ショウジョウバエ遺伝学的解析から同定され、体の前後軸に沿った体節パターンに関わる遺伝子産物の総称。現在、哺乳動物ではおよそ20遺伝子( Phc2、 Mel18、 Ring1b、 Cbx2など)がポリコムタンパク質群に分類されている。ポリコムタンパク質群は、それぞれアミノ酸配列構造が異なるため、独自の機能を持つ。 - 2.自己重合活性
この場合は、Phc2タンパク質の進化的に保存されたアミノ酸配列構造(SAMドメインと呼ばれる)がレゴブロックのように頭尾結合して、重合(数珠つなぎのように繋がる)する特性をいう。 - 3.Phc2遺伝子点変異
Phc2遺伝子のDNA配列上の1塩基置換のこと。アミノ酸レベルで、Phc2タンパク質の307番目ロイシン(CTA)がアルギニン(CGA)に変換するように1塩基を置換した。 - 4.クロマチン
真核細胞内におけるDNAとタンパク質(主にヒストン)の複合体のこと。クロマチンの高次構造は、遺伝子発現のオン・オフに深く関わっている。 - 5.Hox遺伝子
動物の胚発生初期において体軸に沿った体節パターンの決定に関与する分化関連遺伝子群。ヒトやマウスでは、 Hoxa、 Hoxb、 Hoxc、および Hoxdという4つのクラスターを編成し、それぞれ異なる染色体に存在する。各クラスターは10前後の Hox遺伝子で構成される( 図3参照)。 - 6.Cdkn2a遺伝子
主要がん抑制遺伝子の1つで、細胞増殖停止、老化、細胞死を誘導する。さまざまながん細胞において Cdkn2aの欠損や不活性化が確認されている。不活性化の要因としては、ポリコムタンパク質群の過剰発現が示されている。 - 7.早期細胞老化
規定の寿命(細胞分裂回数で表される)に到達することなく、細胞が不可逆的に分裂停止する現象。
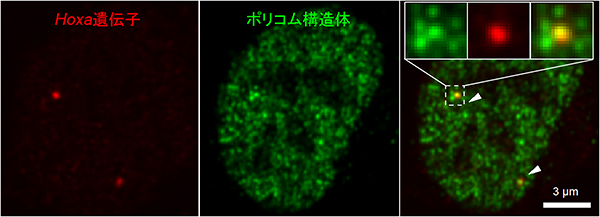
図1 ポリコム構造体
マウス胚由来細胞(胚性繊維芽細胞)の核内に存在するポリコムタンパク質を蛍光タンパク質(GFP)で表している。
- 左: 不活性化状態にあるHoxa遺伝子領域の位置(2つの赤点)。
- 中: 斑点状のポリコム構造体(緑)。
- 右: 左と中を重ね合わせた画像。一部のポリコム構造体(矢じり)は、Hoxa遺伝子領域上に存在している。
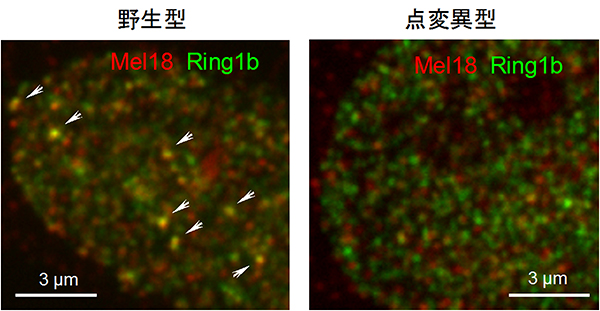
図2 Phc2遺伝子点変異マウス由来の細胞でのポリコム構造体の消失
- 左: 野生型マウスでは、ポリコム構造体(矢じり)がポリコムタンパク質群のMel18タンパク質(赤)とRing1bタンパク質(緑)の共局在によって検出される。
- 右: Phc2タンパク質が重合不全となる点変異マウス(点変異型)の細胞では、Mel18タンパク質とRing1bタンパク質の共局在が確認されない。つまり、ポリコム構造体が消失している。
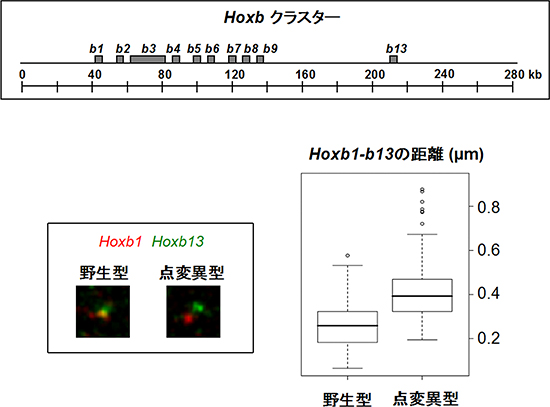
図3 ポリコム構造体形成不全によるHoxb遺伝子群領域のクロマチン構造の緩み
- 上:Hoxb遺伝子クラスターの構造。
- 左下:細胞核内におけるHoxb遺伝子クラスターにあるHoxb1とHoxb13の位置。野生型マウスではHoxb1(赤)とHoxb13(緑)は近接しているが、Phc2遺伝子点変異マウス(点変異型)では離れている。つまり、クロマチン構造が緩んでいる。
- 右下:Hoxb1とHoxb13の2点間距離。
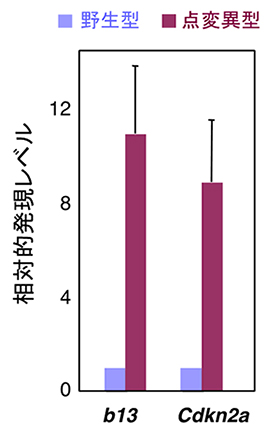
図4 ポリコム構造体の形成不全によって発現が上昇した遺伝子
点変異マウス(点変異型)の細胞では、野生型マウスと比較してポリコム複合体の制御下にあるHoxb13およびCdkn2aの発現が約10倍に上昇していた。
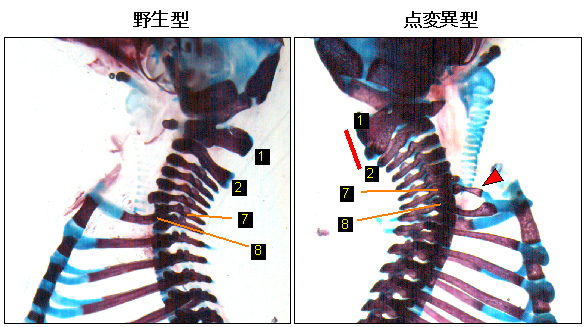
図5 点変異型マウスで起こった体節の運命変換
- 左: 野生型マウスの脊椎は、上から数えて7番目までが頸椎となる。8番目からは肋骨のある胸椎となる。
- 右: 点変異マウス(点変異型)では、第7頸椎が第1胸椎へと運命変換し、肋骨を持つようになる(赤矢じり)。また、第1頸椎も第2頸椎の特性を持ってしまっている(赤太線)。
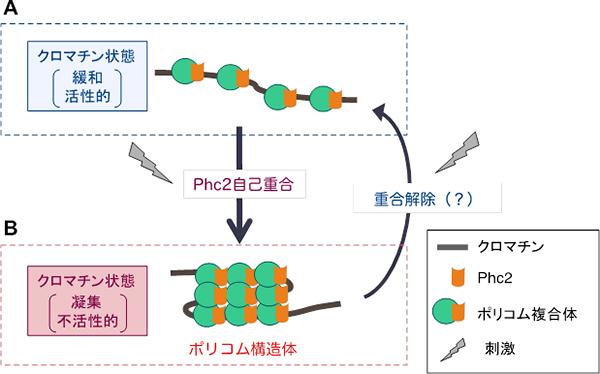
図6 Phc2タンパク質自己重合を介したポリコム複合体による遺伝子発現制御モデル
- A: ポリコム複合体が標的遺伝子座に存在するだけでは不活性化できない。
- B: Phc2タンパク質自己重合がポリコム複合体の重合を引き起こし、ポリコム構造体を形成する。その結果として、クロマチンが凝集し、遺伝子の発現が抑えられる。ポリコム複合体の重合は何らかの刺激によって制御されることで、遺伝子発現のオン・オフが切り換わる可能性がある。