要旨
理化学研究所(理研)環境資源科学研究センター統合メタボロミクス研究グループの梅基直行上級研究員、斉藤和季グループディレクター、大阪大学大学院の村中俊哉教授、神戸大学大学院の水谷正治准教授らの共同研究グループは、ジャガイモに含まれる有毒物質であるソラニンなどの「ステロイドグリコアルカロイド(SGA)[1]」の生合成に関わる遺伝子「PGA1」と「PGA2」を同定し、これらの遺伝子発現を抑制するとSGAを作らなくなるとともに、ジャガイモの萌芽を制御できる可能性を発見しました。
ジャガイモは塊茎[2](かいけい)の緑化した皮の周辺と塊茎から出る芽にSGAが高濃度に蓄積されます。SGA含量が少ないと“えぐみ”などの嫌な味の原因となり、SGA含量が多くなると食中毒を引き起こします。そのため、ジャガイモのSGA含量を低く抑えることは、ジャガイモ育種において重要かつ不可欠です。
また、ジャガイモには収穫後数か月間、成長や発生が一時的に停止する「休眠期間」があります。休眠後に萌芽が始まるため、1年以上の長期保存はできません。したがって、萌芽の制御は年間を通して生産を行うジャガイモ加工業にとって大きな課題です。
SGAはコレステロール[3]を出発物質として生合成されることが知られていますが、SGAに至るまでの生成機構は明らかになっていませんでした。今回、共同研究グループが、SGAが多く蓄積される芽と花で多く発現する遺伝子を解析したところ、PGA1とPGA2を発見しました。そして、PGA2はコレステロールを22-ヒドロキシコレステロールに変換する22位水酸化酵素をコードすること、PGA1は22-ヒドロキシコレステロールを22, 26-ジヒドロキシコレステロールに変換する26位水酸化酵素をコードすることを明らかにしました。
また、PGA1とPGA2の発現をそれぞれ抑制した遺伝子組換え植物体から収穫したジャガイモのSGA含量は、どちらの遺伝子を抑制した場合も非遺伝子組換えジャガイモよりも極めて低下していました。さらに、予想に反して遺伝子組換えジャガイモは休眠期間が過ぎても萌芽しませんでした。加えて土に植えると萌芽し始めました。以上の結果は、PGA1とPGA2の遺伝子発現を抑制したり、ゲノム編集[4]で遺伝子を破壊したりすることで毒がなく、かつ萌芽を制御できるジャガイモを育種できる可能性を示しています。
本研究の一部は、生物系特定産業技術研究支援センター・イノベーション創出基礎的研究推進事業ならびに,総合科学技術・イノベーション会議のSIP(戦略的イノベーション創造プログラム)「次世代農林水産業創造技術」などの支援を受けて行いました。本成果は、米国の科学雑誌『Plant Physiology』(8月号)に掲載されるに先立ち、オンライン版(6月15日付け)に掲載されました。
背景
世界4位の生産量を誇る食用作物であるジャガイモはナス科に属する野菜です。食用にできるのは地下にできる茎で塊茎(かいけい)と呼ばれる部分ですが、日光を浴びて緑化した塊茎の皮の周辺と塊茎から出る芽に、「ステロイドグリコアルカロイド(SGA)」と総称される有毒物質が高濃度に蓄積されます。SGA含量が少ないと“えぐみ”などの嫌な味の原因となり、SGA含量が多くなると食中毒を引き起こします。
管理を誤るとSGAが蓄積されるため、ジャガイモは潜在的に危険な食品といえます。私たちが安心、安全に食している背景には、品種作成(育種)から栽培、収穫後の管理、流通、貯蔵、販売、消費者によって調理されるまで、適切に管理され、そのために相応のコストをかけているという現状があります。
例えば、ジャガイモの品種の一つ、メークインは日光にさらすと、簡単に食中毒の原因となるレベルにまでSGAが蓄積されます。そこまで蓄積されていなくても、ジャガイモに“えぐみ”を感じた経験がある人は多いと思います。そのため、ジャガイモのSGA含量を低く抑えることは、ジャガイモ育種や“食の安全”において重要かつ不可欠です。
ジャガイモに蓄積する主なSGAはソラニン、チャコニンという物質です(図1)。梅基直行上級研究員らは2014年に、SGAがコレステロールを出発物質として生合成されることを明らかにしました注1)。しかし、その後の詳しい生成機構は分かっていませんでした。
また、米や小麦などの穀類とは異なり、ジャガイモには「休眠期間」があります。その名の通り、収穫後数か月間、成長や発生が一時的に停止する“眠り”につきます。休眠後に萌芽が始まるため、1年以上の長期保存はできません。萌芽を制御することはSGAとは別のジャガイモ特有の課題でした。
注1)2014年9月13日プレスリリース「ジャガイモの有毒アルカロイド生合成酵素遺伝子を同定」
研究手法と成果
ステロイド化合物の一種であるコレステロールからSGAへの生合成過程では、少なくとも3カ所で水酸化(ヒドロキシル化、-OHの付加反応)を受けると想定されていました。また、植物ホルモンであるステロイド化合物の酸化過程の代謝反応には、シトクロムP450酸化酵素[5]が関わっていることが知られています。
今回、共同研究グループはSGAが多く蓄積される芽や花で多く発現する遺伝子を解析しました。その結果、シトクロムP450酸化酵素のファミリーをコードする遺伝子「PGA1」と「PGA2」を発見しました。次に、RNA干渉法[6]でPGA1とPGA2の発現をそれぞれ抑制した遺伝子組換え植物体を作り出し、ジャガイモを収穫しました。そのSGA含量を調べたところ、どちらの遺伝子を抑制した場合も、遺伝子組換えを行っていないジャガイモよりも極めてSGA含量が低いことが分かりました。さらに、光を照射してもSGA含量は増加しませんでした。
また、PGA1とPGA2の遺伝子発現を抑制したそれぞれの植物体の精密なアルカロイド・ステロイド分析と、昆虫細胞を用いた異種タンパク質発現系における酵素活性を測定しました。その結果、PGA2はコレステロールを22-ヒドロキシコレステロールに変換する22位水酸化酵素をコードすること、PGA1は22-ヒドロキシコレステロールをさらに22, 26-ジヒドロキシコレステロールに変換する26位水酸化酵素をコードすることが明らかになりました(図2)。
2014年に梅基直行上級研究員らは、コレステロール合成酵素遺伝子である「SSR2遺伝子」の発現を抑制または破壊したジャガイモと遺伝子組み換えなどを行っていない野生型のジャガイモでは生育に差がみられなかったことから、SGA自体は生育に必要ではないことを発見しました注1)。今回、PGA1とPGA2の発現をそれぞれ抑制した場合、同様にジャガイモの収量には差がみられませんでした。しかし、収穫したジャガイモは休眠期間を経過した後、何も処理をしない限り、暗所で4℃で保存しても20℃で保存しても萌芽しないことが分かりました(図3)。ところが、萌芽しないジャガイモを土に植えると、萌芽し始めました。この現象は4℃で3年間保存した後でも再現できました。
これは、ジャガイモではこれまでみられたことのない現象です。SGAの生合成経路の変化により、萌芽を進めるどこかの段階に何らかの攪乱が起き、中断していることが予想できますが今のところ原因は分かっていません。
今後の期待
本成果により、今後、PGA1やPGA2遺伝子を標的としたゲノム編集などによる遺伝子破壊の手法を利用することで、SGA含量を低く抑え、かつ萌芽を制御できるジャガイモの育種が可能となると期待できます。
原論文情報
- Naoyuki Umemoto, Masaru Nakayasu, Kiyoshi Ohyama, Mari Yotsu-Yamashita, Masaharu Mizutani, Hikaru Seki, Kazuki Saito, and Toshiya Muranaka, "Two Cytochrome P450 Monooxygenases Catalyze Early Hydroxylation Steps in the Potato Steroid Glycoalkaloid Biosynthetic Pathway", Plant Physiology, doi: 10.1104/pp.16.00137
発表者
理化学研究所
環境資源科学研究センター 統合メタボロミクス研究グループ
上級研究員 梅基 直行(うめもと なおゆき)
グループディレクター 斉藤 和季(さいとう かずき)
大阪大学大学院工学研究科生命先端工学専攻
教授 村中 俊哉(むらなか としや)
神戸大学大学院農学研究科生命機能科学専攻
准教授 水谷 正治(みずたに まさはる)
 梅基 直行
梅基 直行
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
大阪大学工学研究科総務課評価・広報係
Tel: 06-6879-7231 / Fax: 06-6879-7210
kou-soumu-hyoukakouhou [at] office.osaka-u.ac.jp(※[at]は@に置き換えてください。)
神戸大学総務部広報課
Tel: 078-803-6696 / Fax: 078-803-5088
ppr-kouhoushitsu [at] office.kobe-u.ac.jp(※[at]は@に置き換えてください。)
補足説明
- 1.ステロイドグリコアルカロイド(SGA)
窒素原子を含むステロイドの配糖体。ナス科植物などで生成蓄積されることが知られている。ジャガイモではソラニン、チャコニンが主なSGAとして知られ、ジャガイモ食中毒の原因物質とされている。SGAはSteroid Glycoalkaloidの略 - 2.塊茎
地下茎の一部が養分を蓄積して塊状に大きくなったもの。多年生の草本植物で地上部が毎年枯れるものにはこの形で冬を越すものがある。ジャガイモやコンニャクイモ、クワイなど、食用になるものが多い。植物学的には休眠すること、栄養器官での維管束の配列の違いなどから根と区別できる。 - 3.コレステロール
ステロイドの一種。細胞を構成する脂質成分として脳、神経、胆汁、血液など動物組織一般にみられる。植物では限られた植物にだけ多く含まれる。ナス科植物の場合、総ステロイドの10%程度がコレステロールである。 - 4.ゲノム編集
部位特異的なDNAへの結合を設計できる人工ヌクレアーゼを利用することで、標的遺伝子を改変する技術。ZFN、TALEN、CRISPR/Cas9などの方法があり、これまで遺伝子破壊が困難であった生物でも自由に遺伝子破壊ができるようになり、急速に普及しつつある。 - 5.シトクロムP450酸化酵素
酸化還元酵素ファミリーに属する酵素の総称。さまざまな基質を酸化し、多くの役割を果たす。肝臓で解毒を行う酵素であるとともに、ステロイドホルモンの生合成、脂肪酸の代謝や植物の二次代謝など、生物の正常活動に必要なさまざまな反応に関与している。 - 6.RNA干渉法
標的遺伝子配列の一部に相当する二本鎖RNAによって、標的遺伝子の発現を抑制する手法。
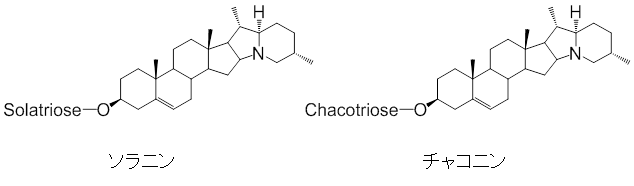
図1 ステロイドグリコアルカロイド(SGA)のソラニンとチャコニンの構造
ステロイドグリコアルカロイド(SGA)は、窒素原子を含むステロイドの配糖体である。ナス科植物には有毒なSGAが含まれる場合がある。代表的なものはジャガイモのソラニンとチャコニンで、芽や緑化した皮の近くに比較的多く含まれる。構造中のSolatrioseとChacotrioseは異なる三つの糖を表している。
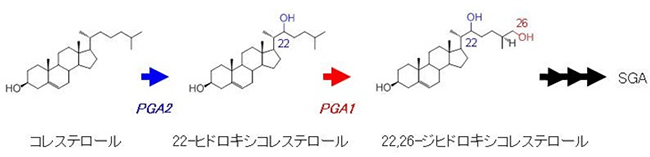
図2 同定したSGAの生合成経路と遺伝子の関係
コレステロールは、22位水酸化酵素によって22位に水酸基が導入され、22-ヒドロキシコレステロールに変換される。この反応の22位水酸化酵素をコードしているのがPGA2である。次に22-ヒドロキシコレステロールの26位が26位水酸化酵素によって水酸化され、22, 26-ジヒドロキシコレステロールになる。この反応の26位水酸化酵素をコードしているのがPGA1である。

図3 遺伝子組換えジャガイモの休眠後の様子
右の2個は、PGA1の発現を抑制した遺伝子組換えジャガイモ。3か月の休眠の後、20℃の暗所に7週間置いたが萌芽は始まらなかった。
左の2個は遺伝子組換えを行っていない野生型のジャガイモ。休眠後、右の遺伝子組換えジャガイモと同じ条件に置いたところ萌芽した。