2025年3月21日
理化学研究所
東京大学大学院理学系研究科
科学技術振興機構(JST)
発現量が進化しやすい遺伝子を細菌で発見
-偏りがある生物進化の予測と制御に期待-
理化学研究所(理研)生命機能科学研究センター 多階層生命動態研究チームの古澤 力 チームリーダー(東京大学 大学院理学系研究科附属生物普遍性研究機構 教授)と東京大学 大学院理学系研究科附属生物普遍性研究機構の津留 三良 特任助教の共同研究チームは、進化実験[1]で得た大腸菌の遺伝子の発現量[2]を網羅的に解析し、遺伝子変異や環境変化で生じる細胞状態の変化に対して、共通して発現量が進化しやすい遺伝子を明らかにしました。
本成果は、細菌の抗生物質耐性獲得進化[3]などで見られるような、発現量が高速に進化するメカニズムの解明への貢献に加え、生物進化の制御によるバイオテクノロジー分野への応用が期待されます。
生物は、個体差や種差などの表現型(生物の遺伝子型が形質として表れたもの)において幅広い多様性を示しますが、これらの多様性は完全にランダムに生じるのではなく、一定の方向性があることが知られています。
今回、共同研究チームは、多様な変異が蓄積しやすい実験条件で大腸菌を進化させ、その遺伝子発現量を解析することにより、発現量が進化しやすい遺伝子の発見に成功しました。さらに、これらの遺伝子は、特定の制御タンパク質(転写因子[4])による発現制御を受けており、遺伝子変異や環境変化の影響を受けやすい制御構造を持つことを突き止めました。本研究の成果は、生物の進化しやすさをつかさどる分子機構に迫るとともに、なぜ生物が特定の状態に進化しやすいかというダーウィン以来の進化の謎に答える可能性を有しています。
本研究は、科学雑誌『Nature Communications』オンライン版(3月21日付:日本時間3月21日)に掲載されました。
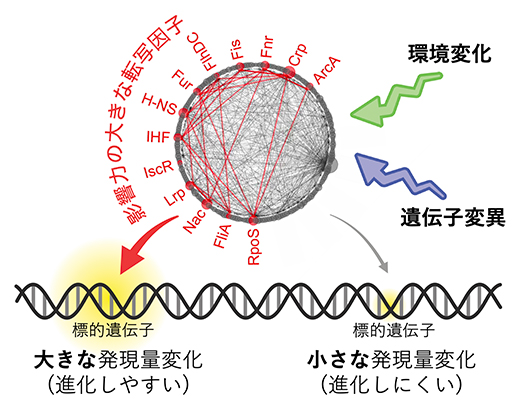
発現量が大きくて進化しやすい遺伝子と、それらを制御する転写因子
背景
生物の形態や機能などの状態は、環境変化や遺伝子変異など、複数の要因によってさまざまに変化し、一世代内で生じる環境変化に対して発現量がより大きく(あるいは小さく)変化できるよう進化します。結果として現在われわれが目にする生物の特筆すべき多様性をつくり上げています。しかしながら、生物の状態変化は実は一様ではなく、特定の状態変化が他よりも高頻度に生じること(偏り)が知られています。こうした偏りは、自然淘汰[5]とは独立に、生物進化を加速・停滞させる効果を持つため、生物進化を統合的に理解する上で重要な現象として注目されてきました。
近年、遺伝子発現を網羅的に解析する技術の発展により、状態変化の偏りを遺伝子レベルで計測し、偏りを生み出す分子機構に迫ろうとする試みが世界的に行われ始めています。その結果、遺伝子ごとに発現量の変化のしやすさが異なることが示唆されてきました。しかしながら、異なる変化の要因(環境変化と遺伝子変異)を厳密に切り分けた上で、多様な変異が蓄積しやすい実験データを取得することは一般的に困難です。そのため、どの遺伝子の発現量がどの要因に対して変化しやすいか、またなぜ変化しやすいかについては、ほとんど明らかにされていません。
これらの課題を解決するため、共同研究チームは、大腸菌を用いた遺伝子発現解析に着目しました。大腸菌は、これまでの研究の蓄積によって、遺伝子ごとに発現量を制御する転写因子がよく分かっており、発現量の変化のしやすさをつかさどる分子機構を探る上で理想的なモデル生物です。また、さまざまな環境条件で培養して得られた遺伝子の発現量プロファイル(遺伝子ごと、培養条件ごとの発現量データ)が公開されており、これらを解析することで、どの遺伝子が環境変化に対してどの程度発現量が変化しやすいかを調べることができます。これらの既存データの解析に加え、今回、共同研究チームは、多様な変異が蓄積しやすい実験条件で大腸菌を進化させ、その遺伝子発現量を解析することにより、環境変化だけでなく、遺伝子変異に対しても発現量が変化しやすく、進化しやすい遺伝子の発見に挑みました。
研究手法と成果
共同研究チームはまず、大腸菌を寒天培地上で培養して得られる単一細胞由来の細胞塊(コロニー)を無作為に選び、植え継ぐことによって、多様な突然変異を蓄積した遺伝子変異株を作製しました。この進化実験で用いた大腸菌株はDNA複製の誤りが起きやすい性質があります。また、植え継ぎ時には1個体(1細胞)ずつに分けられて生存競争の相手(同種他個体)と隔離されるため、個体数が大きな集団では淘汰されてしまうような遺伝子変異株が淘汰されません。結果として、植え継ぎの際に枝分かれする進化系列ごとに、それぞれ独立にさまざまな変異を蓄積することが、津留特任助教らの先行研究注)によって確かめられています。
このようにして得たさまざまな遺伝子変異株を、環境要因の交絡(影響)を防ぐために一定の環境条件で培養し、遺伝子ごとの発現量を計測しました(図1a)。個々の遺伝子の発現量は、ゲノム上に蓄積した多様な遺伝子変異の影響で、遺伝子変異株間で違いが生じ得ます。そこで、個々の遺伝子で見られた遺伝子変異株間の発現量の違い(分散)に基づいて、「遺伝子変異による発現量変化の大きさ(DMmut)」を定量しました。
次に、上記の実験とは逆に、同じ大腸菌株が異なる環境に置かれたときの遺伝子発現量の変化を調べるため、公開データベースから、同一の遺伝情報を持つ大腸菌をさまざまな環境条件で培養して得られた発現量プロファイルを取得しました。これらのプロファイルでは、遺伝子変異の要因の影響が除かれており、個々の遺伝子の発現量の違いは、環境変化のみに依存して生じ得ます。上記と同様の解析を行うことで、遺伝子ごとに「環境変化による発現量変化の大きさ(DMenv)」を定量しました。
これら2種の発現量変化の大きさの関係を調べたところ、環境変化に対して遺伝子変異を伴わずに発現量が大きく変化しやすい遺伝子は、遺伝子変異に対しても発現量が変化しやすいことが明らかとなりました(図1b)。これらの遺伝子は、細胞の運動やアミノ酸代謝に関連する機能を持つ遺伝子に多く見られました。また、これらの遺伝子は本研究の進化実験だけに特異的なものではなく、抗生物質耐性獲得進化や自然界での進化においても、発現量が変化しやすい遺伝子であることが分かりました。一方で、細胞壁の構築や維持に関わる遺伝子群は発現量の変化が少なく、環境変化や遺伝子変異の影響を受けにくい頑健さがありました。これらの結果は、「遺伝子変異によらない表現型の揺らぎと、遺伝子変異を伴う進化の方向性には相関がある」という津留特任助教らの先行研究による理論予測注)を裏付けるものであり、異なる要因による発現量の変化のしやすさが、共通の分子メカニズムによって支配されていることを示唆しています。
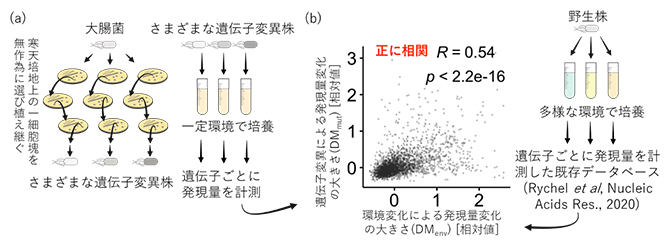
図1 実験室で進化させた大腸菌の遺伝子発現解析と得られた結果の概要
(a)大腸菌を用いた進化実験の概要。寒天培地上に生じた細胞塊(コロニー)を無作為に選び、植え継ぐことで得た遺伝子変異株を、一定の環境で培養し、遺伝子ごとに発現量を計測した。(b)遺伝子変異による発現量変化の大きさ(DMmut)と環境変化によるそれ(DMenv)の正相関(内挿はスピアマンの順位相関係数R(-1から+1の値を取り、絶対値が大きいほど相関が強い)とp値(有意確率))。前者は本研究から、後者は既存のデータベース(Rychelら、Nucleic Acids Res., 2020)を用いて得られた。正相関は、環境変化に対して発現量が変化しやすい遺伝子は、遺伝子変異に対しても発現量が変化しやすいことを意味している。
発現量の変化のしやすさを左右する分子メカニズムを探索するため、共同研究チームは、発現量変化の大きな遺伝子がどのような転写因子によって発現制御を受けているかを調べました。これまでの知見から、大腸菌では、転写因子とそれによって直接発現制御を受ける標的遺伝子群の組み合わせがよく分かっています。この情報に基づいて、変化の要因(環境変化と遺伝子変異)ごと、転写因子ごとに、その標的遺伝子群が大きな発現量変化を示す傾向にあるか否かを判定しました。その結果、二つの要因に共通する13個の転写因子(影響力の大きな転写因子、TRcom)を発見しました(図2)。TRcomによって直接発現制御を受ける標的遺伝子群は、環境変化に対しても遺伝子変異に対しても発現量が変化しやすく、進化しやすいことを意味しています。これらの結果は、異なる変化の要因間に共通する分子メカニズムの存在を裏付けています。
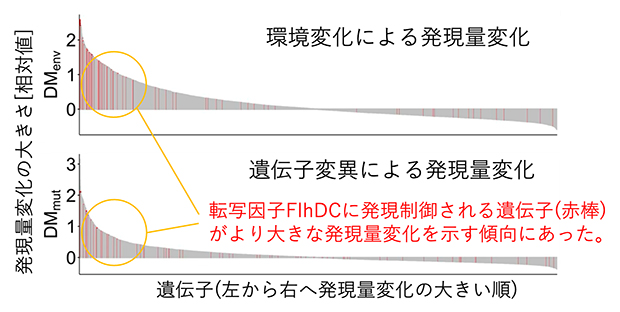
図2 発現量変化の大きな遺伝子を制御する転写因子(TRcom)
影響力の大きな転写因子(TRcom)の検出例(上:環境変化による発現量変化、下:遺伝子変異による発現量変化)。全遺伝子を発現量変化の大きさ順(左から右へ降順)に並べ、注目する転写因子(例では、転写因子FlhDC)によって直接制御される標的遺伝子群の出現位置が赤棒で示されている。無作為で一様な出現を仮定した場合と比較し、統計的に有意に出現位置が偏っているかを調べることで、大きな発現量変化を誘導させやすい転写因子を発見できる。
TRcomがどのように「発現量が大きく変化しやすい遺伝子」を制御しているのかを突き止めるため、転写因子間の転写制御ネットワーク[6]構造を調べました。その結果、TRcomは、遺伝子変異や環境変化によって生じるさまざまな細胞状態の変化を感知し、その影響を複数の標的遺伝子の発現量変化に伝播(でんぱ)しやすい制御構造を持つことが分かりました。生物は、さまざまな細胞状態の変化を感知するセンサーの役割を担うタンパク質を多数保有しています。大腸菌もこうした分子機構を保有しており、センサーを兼ねた転写因子(SR)が多数同定されています。今回発見したTRcomにもSRが含まれており(8/13個)、さらに、TRcomのほとんど(12/13個)が、約30個の異なるSRと強連結成分[7]を形成していることが分かりました(図3)。転写制御ネットワークにおける強連結成分は、互いに直接あるいは間接的に制御し合う転写因子の集合のことであり、TRcomの多くが、多数のSRが感知した細胞状態の変化を受信できる制御構造を持つことを意味しています。さらに、TRcom間にも、統計的に有意に多くの発現制御が張り巡らされていることを見いだしました。こうした制御構造は、TRcomによって発現制御を受ける標的遺伝子が、さまざまな環境変化や遺伝子変異に対して、なぜ高頻度に発現量が変化しやすいかを説明するものであり、発現量の進化のしやすさをつかさどる分子機構に深く関わっていると予想されます。
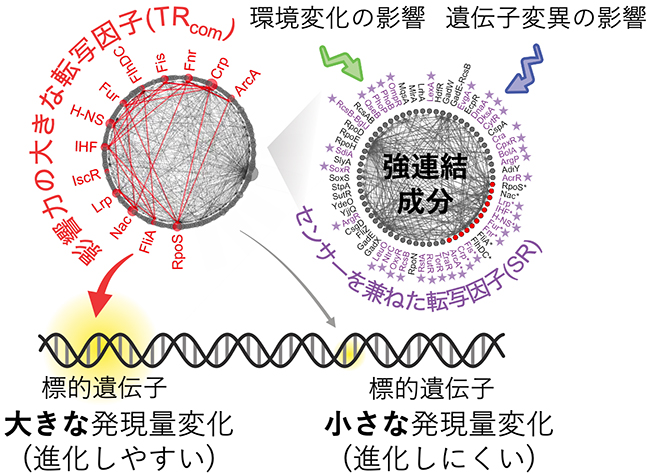
図3 細胞状態の変化を感知し、遺伝子の発現量変化に伝播する転写制御ネットワーク
大腸菌の転写制御ネットワーク。影響力の大きな転写因子(TRcom)が赤丸でラベルされ、線は転写因子間の制御を表す。右は、転写制御ネットワークの中の最大強連結成分。環境変化や遺伝子変異の影響を感知する既知の仕組みを持つ転写因子(センサーを兼ねた転写因子、SR)が紫星印でラベルされている。強連結成分内の転写因子は、互いに直接あるいは間接的な発現制御を受けている。
- 注)Tsuru, S., Ishizawa, Y., Shibai, A., Takahashi, Y., Motooka, D., Nakamura, S. and Yomo, T. (2015), Genomic confirmation of nutrient-dependent mutability of mutators in Escherichia coli. Genes Cells, 20: 972-981. doi 10.1111/gtc.12300
今後の期待
本研究では、進化実験で得た大腸菌の遺伝子発現量変化を網羅的に解析し、発現量が進化しやすい遺伝子を明らかにしました。これらの遺伝子は、抗生物質耐性獲得進化を含むさまざまな進化においても発現量を進化させやすい性質を示しています。今後、これらの遺伝子の発現量進化がどのように細菌の環境適応に役立てられているかを調べることで、院内感染などで問題となる抗生物質耐性菌の進化が高速に生じるメカニズムの解明が期待されます。
また、本研究では、発現量を進化させやすい遺伝子が、特定の転写因子による発現制御を受けていることを発見しました。さらに、これらの転写因子が、遺伝子変異や環境変化によって生じるさまざまな細胞状態の変化を感知し、その影響を複数の標的遺伝子の発現量変化に伝播させやすい制御構造を持つことを突き止めました。今後、これらの制御構造を人為的に改変あるいは応用し、その効果を確認することで、発現量進化の制御による有用細菌の育種の効率化など、バイオテクノロジー分野への応用が期待されます。
補足説明
- 1.進化実験
複雑な自然環境で進化した生物種を単に採取して調べるのではなく、実験室内のよく制御された環境下で生物を進化させる実験のこと。 - 2.発現量
DNAにコードされている遺伝子の情報は、発現(転写・翻訳)と呼ばれる化学反応によって読み出され、RNAやタンパク質などの遺伝子産物が合成される。本研究では各遺伝子から発現されたRNAの細胞内の存在量をその遺伝子の発現量としている。 - 3.抗生物質耐性獲得進化
細菌などの増殖や生理機能を阻害する化学物質は抗生物質と呼ばれ、感染症の治療などに用いられる。初期には抗生物質への耐性が弱い細菌が、耐性を強める進化のことを抗生物質耐性獲得進化と呼ぶ。抗生物質が効かなくなるため、抗生物質耐性獲得進化は医学上の大きな問題となっている。 - 4.転写因子
DNAに結合して遺伝子からRNAへの発現(特に転写)を制御(促進あるいは抑制)する機能を持ったタンパク質のこと。本研究では、転写因子によって制御を受ける遺伝子を標的遺伝子と呼ぶ。 - 5.自然淘汰
生物進化において、環境への適応能力の低い個体が、適応能力の高い個体との生存競争に負けて絶滅していくこと。 - 6.転写制御ネットワーク
転写因子が自己あるいは別の転写因子の遺伝子の発現を制御することで形成されるネットワークのこと。ノード(ネットワークを構成する要素)を転写因子、リンク(要素同士をつなぐ線)を転写因子間の制御として図示されることが多い。 - 7.強連結成分
リンク(連結)に向きを持つネットワークは有向グラフと呼ばれる(転写制御では、ある転写因子から別の転写因子への制御の向きを持つ)。強連結成分とは、有向グラフにおいて、リンクの向きに従って互いに行き来が可能なノードの集合のこと。転写制御ネットワークでは、制御関係に従って直接あるいは間接的に制御し合う転写因子の集合を意味する。
研究支援
本研究は、理化学研究所運営費交付金(生命機能科学研究)で実施し、日本学術振興会(JSPS)科学研究費助成事業挑戦的研究(萌芽)「発現量の進化しやすさを調節する非コーディング領域の探索(研究代表者:津留三良)」、同新学術領域研究(研究領域提案型)「進化の制約と方向性~微生物から多細胞生物までを貫く表現型進化原理の解明~(領域代表者:倉谷滋)」、同国際共同研究加速基金(国際先導研究)「太陽系に広がる惑星環境における前生命化学進化と分子システム誕生の多様性の解明(研究代表者:廣瀬敬)」、科学技術振興機構(JST)戦略的創造研究推進事業ERATO「深津共生進化機構プロジェクト(研究総括:深津武馬、JPMJER1902)」などによる助成を受けて行われました。
原論文情報
- Saburo Tsuru and Chikara Furusawa, "Genetic properties underlying transcriptional variability across different perturbations", Nature Communications, 10.1038/s41467-025-57642-8
発表者
理化学研究所
生命機能科学研究センター 多階層生命動態研究チーム
チームリーダー 古澤 力(フルサワ・チカラ)
(東京大学 大学院理学系研究科附属生物普遍性研究機構 教授)
東京大学 大学院理学系研究科附属生物普遍性研究機構
特任助教 津留 三良(ツル・サブロウ)
 古澤 力
古澤 力
 津留 三良
津留 三良
JST事業に関すること
科学技術振興機構 研究プロジェクト推進部 ICT/ライフイノベーショングループ
今林 文枝(イマバヤシ・フミエ)
Tel:03-3512-3528
Email:eratowww@jst.go.jp
報道担当
理化学研究所 広報室 報道担当
お問い合わせフォーム
東京大学 大学院理学系研究科・理学部 広報室
Email: media.s@gs.mail.u-tokyo.ac.jp
科学技術振興機構 広報課
Tel: 03-5214-8404
Email: jstkoho@jst.go.jp