2016年10月27日
理化学研究所
徳島大学
医薬基盤・健康・栄養研究所
1分子の構造変化による細胞集団運動の制御
-異分野領域融合研究により高次細胞機能の謎を解く-
要旨
理化学研究所(理研)光量子工学研究領域画像情報処理研究チームの横田秀夫チームリーダー、吉澤信上級研究員、徳島大学大学院医歯薬学研究部の佐々木卓也教授、坂根亜由子助教、医薬基盤・健康・栄養研究所の水口賢司プロジェクトリーダーらの共同研究グループ※は、JRABというタンパク質の1分子の構造変化に着目し、細胞集団運動の制御機構を解明しました。
複数の細胞からなる細胞集団の運動は、胎生期の組織・器官形成の過程だけでなく、創傷治癒やがん転移などでも広くみられる現象です。細胞集団の動きは複雑なことから、これまでは多数の分子が関与すると予想されていました。しかし、共同研究グループは、これまでの研究により低分子量Gタンパク質[1]Rab13の標的タンパク質として発見されたJRABが細胞集団運動において重要な役割を果していると考え、JRABというたった1分子の構造変化に着目して、細胞集団運動の制御機構を解明しようと試みました。
まず、バイオインフォマティクス(生命情報学)と生化学的実験を組み合わせることで、JRABのRab13との結合による構造変化モデルを示しました。またJRABの野生型や構造変異体(open formとclosed form)を発現させた3種類の細胞集団の動きの異なった特徴を、ライブイメージング像の時空間ボリュームレンダリング[2]による解析で抽出・可視化に成功しました。さらに、オプティカルフロー[3]と主成分分析[4]を組み合わせ、画像の輝度変化に強い手法を開発し、従来法では困難だった細胞集団の動きの計算と膨大な情報の定量的な解析を実現しました。その結果、構造を自由に変化できる野生型のJRABは、open formやclosed form変異体と比較して最も効率の良い細胞集団の動きを可能にしていることを証明しました。また、開発したバイオメカニクス(生体力学)の手法を用いた解析ではclosed form のJRABが細胞集団の先頭の一部で集団を引っ張るのに必要な力を生み出していることが分かりました。
今回、生化学、細胞生物学、コンピュータサイエンス、バイオインフォマティクス、バイオメカニクスといった異分野領域の融合研究によって、細胞集団運動という高次細胞機能の制御をタンパク質1分子の構造変化で説明することができました。本研究は今後、発生異常の病態解明やがん転移機構の解明につながると期待できます。
本研究の一部は、科学研究費補助金新学術領域研究「病態の理解に向けた細胞内物流システムの融合研究」「動く細胞と場のクロストークによる秩序の生成」の支援を受けて実施しました。成果は、米国の科学雑誌『Molecular Biology of the Cell』(10月15日号)に掲載されました。
※共同研究グループ
理化学研究所 光量子工学研究領域
エクストリームフォトニクス研究グループ 画像情報処理研究チーム
チームリーダー 横田 秀夫(よこた ひでお)
上級研究員 吉澤 信(よしざわ しん)
テクニカルスタッフⅠ 西村 将臣(にしむら まさおみ)
徳島大学
大学院医歯薬学研究部
医科学部門 生化学分野
教授 佐々木 卓也(ささき たくや)
助教 坂根 亜由子(さかね あゆこ)
特定研究部門 光イメージング研究分野
教授 堀川 一樹(ほりかわ かずき)
医薬基盤・健康・栄養研究所 バイオインフォマティクスプロジェクト
プロジェクトリーダー 水口 賢司(みずぐち けんじ)
研究員(研究当時) 土屋 裕子(つちや ゆうこ)(大阪大学蛋白質研究所 助教)
大阪大学大学院基礎工学研究科 生体計測学講座
教授 出口 真次(でぐち しんじ)
北海道大学大学院先端生命科学研究院 細胞ダイナミクス科学研究室
教授 芳賀 永(はが ひさし)
背景
「細胞集団運動」では、多くの細胞が一つの集団としてまとまって動きます。この現象は、発生・再生時の組織・器官形成過程だけでなく、創傷治癒やがんの転移過程などでも広くみられ、その制御機構の理解は医学・生物学的に重要です。細胞集団運動では、細胞集団の先頭の一部の細胞において集団を一方向に牽引するのに必要な強い力が生み出される一方で、後方の大多数の細胞において隣の細胞と離れないようにしっかりと接着を維持しながら細胞間でのコミュニケーションをとっており、これらによって細胞集団として統率のとれた振る舞いが可能になっています。
細胞集団運動の制御因子として、これまでにさまざまな機能分子が報告されてきたことから、多くの分子が複雑に絡み合った制御機構の存在が予想されていました。徳島大学のグループはこれまでに、低分子量Gタンパク質Rab13とその標的タンパク質JRABが、細胞間接着分子の輸送を介して細胞間接着形成を制御することを明らかにしています注1)。さらに、このRab13-JRABシステムが、アクチン細胞骨格[5]の再編成を時空間的に制御することを示していました注2)。また、生化学的解析からJRABは、Rab13との相互作用によってclosed formからopen formへと構造変化を引き起こすことが示唆されています。さらに、open formやclosed formのままで構造変化できない変異体JRABΔCCやJRABΔCTを発現させた細胞の集団運動には異常が認められることから、JRABの構造変化が細胞集団運動において重要な役割を果たしていることが予想されました。そこで、共同研究グループは、JRABというたった1分子の構造変化に着目して、細胞集団運動の制御機構の解明を試みました。
注1)Terai, T. et al.JRAB/MICAL-L2 Is a Junctional Rab13-binding Protein Mediating the Endocytic Recycling of Occludin Mol.Biol.Cell, 17 (5), 2465-2475 (2006)
注2)Sakane, A. et al. Rab13 Small G Protein and Junctional Rab13-binding Protein (JRAB) Orchestrate Actin Cytoskeletal Organization during Epithelial Junctional DevelopmentJ.Biol.Chem., 287(51), 42455-42468 (2012)
研究手法と成果
共同研究グループは、まず、JRABの構造変化の実態を明らかにするため、バイオインフォマティクス(生命情報学)を用いたJRABの構造モデリングを試みました。in silicoの立体構造モデリング[6]の各過程で生化学的実験を行い、得られたモデルの検証・修正を繰り返すことでRab13-JRAB複合体の構造モデルを完成させました。その結果、JRABの分子内結合がRab13との相互作用により解除されて構造変化が引き起こされることを構造学的に証明しました(図1)。
加えて、JRABのN末端側とC末端側[7]の結合・非結合を感知することにより、open formとclosed form間の構造変化を捉えることができる「JRABのFRETプローブ[8]」を用いて、細胞集団内でのJRABの構造の時空間変化を証明しました(図2)。
これまで、JRABの構造変異体JRABΔCC(open formのまま構造変化できない変異体)やJRABΔCT(closed formのまま構造変化できない変異体)を発現させた細胞の集団運動では、各構造変異体に特徴的なライブイメージング像が認められていました。しかし、その複雑な動きから特徴を抽出することは従来法では困難でした。そこで共同研究グループは、各構造変異体に特徴的なライブイメージング像の時空間を三次元画像として、「ボリュームレンダリング」による解析を行いました。
その結果、JRABΔCTを発現させた細胞集団はラインダンスのように直線的に移動し、JRABΔCCを発現させた集団は阿波踊りのように進行方向を種々の方向に分散させるといった、野生型とは異なる特徴を抽出・可視化することに成功しました(図3)。
また、細胞移動の定量画像解析では、対象物を認識し移動を追跡することで定量化を行うことが一般的でした。しかし集団で動く細胞の解析では、個々の細胞の蛍光輝度の変化が大きいため、集団内の細胞を個別に認識することは困難でした。そこで共同研究グループは、個々の細胞の動態に注目して細胞集団の動きを捉えるのではなく細胞集団動態そのものを捉えて解析する手法として、オプティカルフローと主成分分析を組み合わせた、画像の輝度変化に強い新たな手法を開発し、細胞集団の動きの自動定量化を実現しました。この手法により得られた20億個以上の移動ベクトルの情報を定量的に解析することで、JRABΔCTとJRABΔCCの中間の運動形態をとり、open formとclosed formを適切に使い分けている野生型のJRABを発現させた細胞集団が、最も効率のよい動きを示すことが明らかになりました。
さらに、細胞が生み出す力を基質にかかるシワとして検出することができる「バイオメカニクス(生体力学)の手法」を用いた解析からは、JRABはその構造によって異なる場所で異なるアクチン細胞骨格の制御を行っており、JRABΔCTは細胞集団の先頭の一部で集団を引っ張るのに必要な力を生み出し、JRABΔCCは力を生み出さずに集団の後方に位置する細胞で安定した細胞間接着を維持していることが明らかになりました(図4)。
最後に、イヌ腎臓尿細管上皮細胞由来のMDCK細胞集団に野生型のJRAB、JRABΔCT、JRABΔCCを発現させてコラーゲンゲル内で三次元培養しました。野生型を発現させた細胞集団では通常通り扁平な一つの管腔が形成されましたが、JRABΔCT、JRABΔCCを発現させた細胞集団では複数の管腔を形成するという異常が認められました。これは、JRABの構造変化が管腔形成に至る細胞集団運動に重要であることを示しています(図5)
今後の期待
細胞集団運動の制御機構は、多数の分子が複雑に絡み合ったものであるという捉え方がこれまでは主流でしたが、本研究ではJRABというたった1分子の構造変化が細胞集団のバランスのとれた効率のよい振る舞いを可能にしているということを細胞レベルで証明しています(図6)。
細胞集団運動におけるシンプルな制御機構モデルは、発生異常の病態解明やがん転移機構の解明につながる可能性があり、医学への貢献も期待できます。今後は、解析を個体レベルへと発展させ、生体内でのJRABの構造変化の生理的意義やJRABの構造変化の破綻によって引き起こされる疾患との関連性の解明を目指します。
原論文情報
- Ayuko Sakane, Shin Yoshizawa, Masaomi Nishimura, Yuko Tsuchiya, Natsuki Matsushita, Kazuhisa Miyake, Kazuki Horikawa, Issei Imoto, Chiharu Mizuguchi, Hiroyuki Saito, Takato Ueno, Sachi Matsushita, Hisashi Haga, Shinji Deguchi, Kenji Mizuguchi, Hideo Yokota & Takuya Sasaki, "Conformational plasticity of JRAB/MICAL-L2 provides ‘law and order’ in collective cell migration", Molecular Biology of the Cell, doi: 10.1091/mbc.E16-05-0332
発表者
理化学研究所
光量子工学研究領域 エクストリームフォトニクス研究グループ 画像情報処理研究チーム
チームリーダー 横田 秀夫(よこた ひでお)
上級研究員 吉澤 信(よしざわ しん)
徳島大学
大学院医歯薬学研究部 医科学部門 生化学分野
教授 佐々木 卓也(ささき たくや)
助教 坂根 亜由子(さかね あゆこ)
医薬基盤・健康・栄養研究所 バイオインフォマティクスプロジェクト
プロジェクトリーダー 水口 賢司(みずぐち けんじ)
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.低分子量Gタンパク質
分子量20-30kDaのGTP結合タンパク質。GDP結合型からGTP結合型への転換により活性型となり、特異的な標的タンパク質に結合して細胞内シグナルを伝達する分子スイッチとして機能する。Ras、Rho、Rab、Ran、Arf/Sar1の5つのグループに分類され、増殖、分化、遺伝子発現、運動、小胞輸送などの細胞機能を制御する。 - 2.ボリュームレンダリング
科学・工学データの可視化に用いられる三次元画像を描画するコンピュータ・グラフィクスの手法。視点と描画面の二次元画素を結ぶ直線に沿って三次元画素の輝度値を伝達関数を用いて畳み込み(重み付き平均)、三次元画像の中身を可視化する。今回理化学研究所画像情報処理研究チームで独自に開発したソフトウェア VoTracer(井尻敬客員研究員)を用いて細胞集団の移動を時空間三次元画像として可視化した。 - 3.オプティカルフロー
物体・背景が変形したり動いたりする時系列の画像データ(監視カメラの映像や天候の衛星動画など)に対して、局所的な移動ベクトル(速度と方向)を推定する計算モデル・方法の総称。従来、細胞集団運動の定量化法では、各細胞に対して細胞質や細胞核の領域抽出を行い、時系列でその重心や形状の変化が解析されてきた。しかし、本研究で対象としているJRABタンパク質の変異体などは、細胞内外の力学的・生化学的な条件により常に変動している。それゆえ、細胞内で特定の形状や位相が決まっていない物質や細胞内小器官では従来の機械学習や幾何学的方法による領域抽出が困難であった。そこで本研究ではオプティカルフローの方法を適応してJRAB変異体の時系列画像を場の移動(局所的な移動の集合)として捉え、統計的に解析した。 - 4.主成分分析
統計における分散の多次元への拡張:共分散行列の固有解析により、データを(最小二乗的に)代表する直交軸とその強度を計算する。本研究では、オプティカルフローで推定した細胞集団の二次元移動ベクトル群を代表する二つの直交する移動方向とその速度を計算した。それにより、異なる変異体での移動方向・速度に特異的な違いを見出した。主成分分析は異常値に対して不安定であることが知られており、本研究では中央値に基づいた二次元頻度表を用いることにより、堅固な計算結果を得た。 - 5.アクチン細胞骨格
アクチンは真核細胞に最も多量に含まれるタンパク質で、細胞骨格を作るタンパク質の一つ。アクチン同士が重合、または脱重合することにより、細胞の形態や運動が制御される。アクチン細胞骨格は、微小管とともに細胞の形態形成・維持に働く。粒状のアクチンモノマーが重合してアクチン線維を形成することにより、細胞の形態の維持・変化や細胞の運動に働く。 - 6.in silicoの立体構造モデリング
これまでに蓄積されたタンパク質の既知の二次構造、三次構造についてのデータベースの情報をもとに、未知のタンパク質の立体構造(三次構造)を予想すること。 - 7.N末端側とC末端側
タンパク質はアミノ酸が鎖状に多数結合してできているが、その鎖の先頭の端(アミノ基側)をN末端側、尾部の端(カルボキシル側)をC末端側と呼ぶ。 - 8.FRETプローブ
FRET はFörster resonance energy transfer(フェルスター共鳴エネルギー移動または蛍光共鳴エネルギー移動)の略称。近接した2個の蛍光分子が存在するとき、一方の蛍光タンパク質(供与体)で吸収された光エネルギーが他方の蛍光タンパク質(受容体)に距離依存的に移動することで、受容体からも蛍光が放射される。今回用いたJRAB指示薬では、N末端側に理研の細胞機能探索技術開発チームの宮脇敦史チームリーダーらが開発したVenus(受容体)が、C末端側にCFP(供与体)が配置されており、JRAB分子の立体構造変化を受容体/供与体の蛍光強度比として検出している。
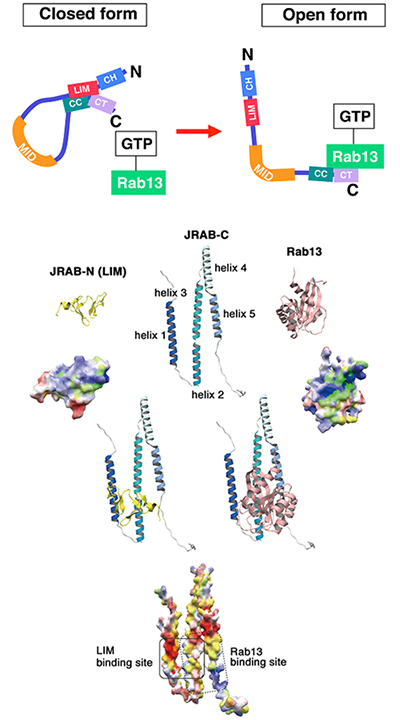
図1 JRABの構造変化とバイオインフォマティクスによるモデリング
生化学とバイオインフォマティクスを組み合わせた解析によってJRABのC末端側(JRAB-C)とN末端側{JRAB-N(LIM)}およびJRABのC末端とRab13の複合体の構造モデルを完成させ、JRABがclosed formからopen formになる過程で、Rab13がC末端に結合するため、競合的にJRABのN末端はC末端から離れることを構造学的に証明した。
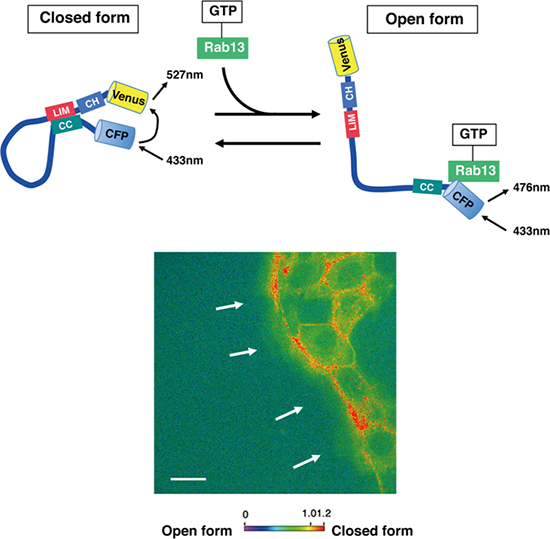
図2 JRABのFRETプローブによる細胞集団内でのJRABの構造変化の検出
上:蛍光タンパク質Venus(黄)をJRABのN末端側に、他の蛍光タンパク質CFP(水色)をC末端側にそれぞれ結合させてN末端側とC末端側の結合・非結合を感知することで、open form(右)とclosed form(左)間の構造変化を捉えることができるJRABのFRETプローブを作製した。
下:FRETプローブを発現させた細胞を用いて、JRABの構造の時空間変化を調べた。写真の赤いシグナルはJRABのN末端側とC末端側の結合、つまり、JRABがclosed formであることを示している。細胞集団の先頭(白矢印)の一部の細胞でJRABはclosed formをとり、後方の大多数の細胞(右側)ではopen formであることが分かる。スケールバーは20マイクロメートル(μm、1μmは1,000分の1mm)。
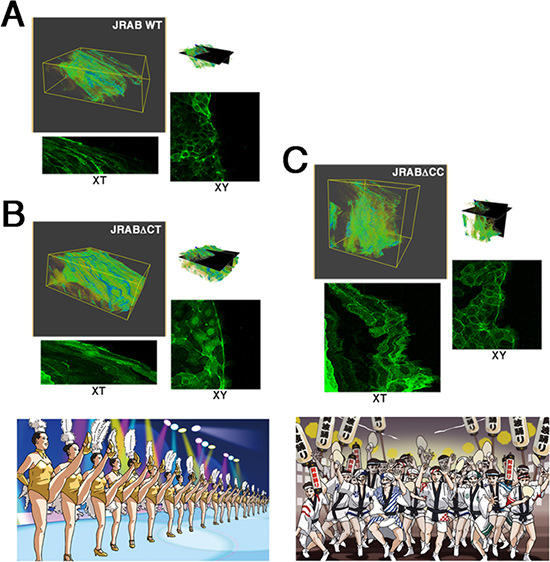
図3 細胞集団運動のライブイメージング像のボリュームレンダリングによる解析
AはJRABの野生型、BはJRABΔCT(closed form変異体)、CはJRABΔCC(open form変異体)をそれぞれ発現させた細胞集団の動きを捉えたライブイメージング像の時空間(XY-Tの三次元データ)ボリュームレンダリングによる解析の結果。Bの細胞集団はラインダンスのように直線的に移動し、Cの細胞集団は、阿波踊りのように進行方向を種々の方向に分散させた。Aの細胞集団は、BとCの中間の運動形態を示した。
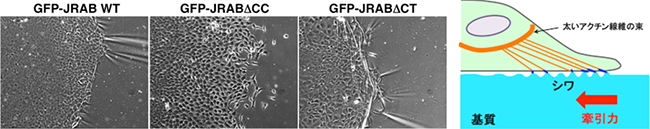
図4 バイオメカニクスを用いた手法による細胞集団内でJRABが生み出す力の検出
画像:細胞が生み出す力をシワとして検出できる基質の上で、野生型のJRAB(左)、open form変異体のJRABΔCC(中央)、closed form変異体のJRABΔCT(右)をそれぞれ発現させた細胞を培養して集団を形成させた。JRABΔCTは細胞集団の先頭(画像の真ん中あたり)で多数のシワを生み出していた(右)。JRABΔCCでは、シワは全く検出されなかった(中央)。野生型では、シワは認められたがJRABΔCTよりは少なく弱いものであった(左)。このことから、closed formのJRABは細胞集団を牽引する力を生み出す役割を担っていると考えられる。
模式図:細胞が生み出す力を基質にかかるシワとして検出することができるバイオメカニクス(生体力学)の手法を表した図。太いアクチン線維の束が、細胞集団を引っ張るのに必要な力(牽引力)を生み出す。
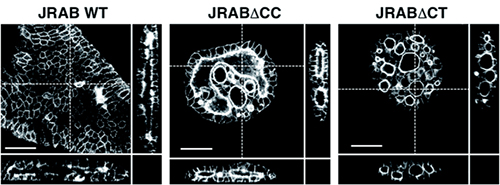
図5 三次元環境下での管腔形成におけるJRABの構造変化の重要性
コラーゲンゲル上で、野生型のJRAB、closed form変異体のJRABΔCT、open form変異体のJRABΔCCをそれぞれ発現させたイヌ腎臓尿細管上皮細胞由来のMDCK細胞を培養し、さらにその上にコラーゲンゲルを重層し、コラーゲンゲル内での管腔形成を観察した。通常は、野生型のJRABを発現させた細胞集団のように扁平な一つの管腔が形成される(左画像)。JRABΔCT(右画像)とJRABΔCC(中央画像)を発現させた細胞集団では、複数の管腔を形成するという異常が認められた。これは、三次元環境下においてもJRABの構造変化が細胞集団運動において重要であることを示している。
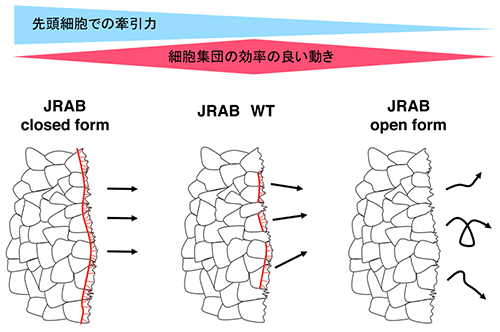
図6 JRABの構造変化と細胞集団運動
closed form変異体のJRABΔCTを発現させた細胞集団は直線的に移動し(左)、open form変異体のJRABΔCCを発現させた集団は進行方向が分散する(右)。しかし、野生型のJRABを発現させた細胞集団は、JRABΔCTとJRABΔCCの中間の運動形態を示し、最も効率のよいバランスのとれた動きが可能となる(中央)。